Расы и штаммы хлебопекарных дрожжей. Различия между расами винных дрожжей. Общая характеристика пивных рас и штаммов
Раздел 3.Производство пива.
Глава 6. Брожение пивного сусла.
Пивные дрожжи .
Строение дрожжевой клетки . Дрожжи - одноклеточные организмы, относящиеся к классу сумчатых грибов. Форма дрожжевых клеток бывает овальной, округлой и эллиптической.
Дрожжевая клетка имеет клеточную стенку 1, под которой располагается цитоплазматическая мембрана. Мембрана обладает избирательной проницаемостью, оказывая влияние на обмен веществ между клеткой и средой.
Например, молекулы аминокислот и глюкозы проникают через мембрану быстрее, чем ионы металлов, которые меньше по размеру. Внутри клетки содержится круглое или овальное ядро 2, окруженное двойной мембраной. Внутри ядра расположено ядрышко. Ядро необходимо для процессов обмена веществ, обеспечивающих рост и
размножение дрожжей.
Основой клетки является цитоплазма 3, представляющая собой вязкую, слегка желтоватую жидкость. Она выполняет многие функции так, например, первая стадия дыхания и спиртовое брожение протекают непосредственно в цитоплазме. Здесь же находятся структурные элементы клетки: вакуоль 4, митохондрии 5, рибосомы 6. Митохондрии- это очень мелкие частицы каплеобразной формы, в которых происходят процессы, связанные с окислительным обменом веществ. Рибосома представляет собой пузырек, окруженный мембраной. Рибосомы являются местом, где происходит синтез белков. Вакуоли- полости, наполненные клеточным соком и отделенные от цитоплазмы вакуолярной мембраной. В них находится метахроматин, обусловливающий рост и размножение дрожжевых клеток. В вакуолях протекают окислительно –восстановительные процессы.
Величина дрожжевых клеток зависит от расы, физиологического состояния дрожжей и состава питательной среды. Прессованные дрожжи содержат около 30% сухих веществ и 70% воды. В сухих веществах дрожжей содержится 90-95% органических веществ и 5-10% неорганических веществ. Среди органических веществ имеются белки и азотсодержащие вещества-54-56%, углеводы-24-40%, жиры-2-4% (к массе сухих веществ). основная часть углеводов представлена гликогеном(запасное вещество), сходным по химстроению с амилопектином крахмала. Среди неорганических веществ около половины фосфорной кислоты и 1/3 калия.
В золе дрожжей содержится (в %): Р 2 О 5 –47-53, К 2 О- 28-40; СаО-0,4 –11,3; Мg О-3,0-7,4;SiO 2 -0,28- 0,73; SiO 3 - 0,09-0,74; Сl –0,1-0,65. Кроме того, в небольшом количестве имеются S , Zn , Mn , Cu , Fe .
Фосфорные соединения имеют важное значение в обмене веществ дрожжевых клеток, так как входят в состав промежуточных веществ спиртового брожения, а калий играет первостепенную роль в построении молекул белков и углеводов. Дрожжи богаты витаминами группы В, содержат эргостерин(провитамин D ) и др. В дрожжах содержатся различные ферментные системы, участвующие в процессах гидролиза и синтеза, а также в процессах брожения и дыхания.
Стадия роста дрожжей . Ростом дрожжей называют увеличение числа их клеток, т.е. – размножение. Дрожжевые клетки при нормальных условиях размножаются почкованием. Материнская клетка образует почку, которая вырастает в дочернюю клетку. При недостатке питательных веществ или при других неблагоприятных условиях внутри клетки образуются перегородки, и клетка распадается по этим перегородкам, образуя споры. В среде с хорошими условиями питания споры прорастают и образуют новые дрожжевые клетки. Пивное сусло содержит все необходимые вещества для размножения клеток, поэтому при сбраживании сусла дрожжи размножаются только почкованием, не образуя спор.
После введения дрожжей в сусло наблюдаются их количественные и качественные изменения. Количество дрожжей Увеличивается в несколько раз, однако их концентрация в диспергированном состоянии вначале увеличивается, достигая максимальной величины, а затем снижается. Размножение дрожжей при сбраживании пивного сусла проходит в несколько этапов. На кривой роста можно выделить несколько этапов. (По вертикали- число дрожжевых клеток, по горизонтали – время.)
 В
начальной фазе, называемой латентной или лаг- фазой (задержка роста), дрожжи
приспосабливаются к новой среде и подготавливаются к размножению. Эту фазу
условно делят на две части: фазу действительного покоя, когда клетки
приспосабливаются к среде,и фазу
постепенного начала размножения. Продолжительность латентной фазы для пивных
дрожжей 1-1,5 сут. В ней клетки увеличиваются в объеме и удлиняются, растет
доля почкующихся клеток.
В
начальной фазе, называемой латентной или лаг- фазой (задержка роста), дрожжи
приспосабливаются к новой среде и подготавливаются к размножению. Эту фазу
условно делят на две части: фазу действительного покоя, когда клетки
приспосабливаются к среде,и фазу
постепенного начала размножения. Продолжительность латентной фазы для пивных
дрожжей 1-1,5 сут. В ней клетки увеличиваются в объеме и удлиняются, растет
доля почкующихся клеток.
При следующей фазе, называемой логарифмической, скорость размножения дрожжей максимальная, все клетки активны и находятся в бродящей среде во взвешенном состоянии.
После логарифмической фазы наступает стационарная фаза, когда размножение клеток замедляется, при этом скорость отмирания и размножения уравновешиваются, в результате чего число живых клеток остается без изменения.
Последняя фаза, называемая фазой затухания, характеризуется снижением активности клеток, что обусловлено уменьшением массы питательных веществ и увеличением количества продуктов обмена. Размножение прекращается, клетки отмирают и оседают на дно бродильного аппарата.
В живой дрожжевой клетке жизнедеятельность поддерживается различными биохимическими процессами, а при её отмирании согласованность этих процессов нарушается и начинается автолиз, т.е. распад клетки под действием собственных ферментов. При этом структура клеток нарушается, повышается активность у одних ферментов и ослабевает у других.
Например, гидролитические ферменты активизируются, а ферменты дыхания и брожения погибают. При автолизе дрожжей происходит распад белковых веществ, углеводородов, жиров, органических фосфорных соединений, образуются низкомолекулярные продукты распада, которые диффундируют через стенки клеток в пиво и изменяют его вкус. При незначительном автолизе появляется слабый дрожжевой привкус, а при сильном автолизе - горький посторонний вкус. Выделяемые при автолизе азотистые вещества могут быть коллоидного помутнения пива.
Расы пивных дрожжей. В пивоваренном производстве используют только культурные дрожжи, которые относятся к семейству сахаромецетацеа (Saccharomycetaceae ) и к роду сахаромицес (Saccharomyces ). Различают дрожжи низового брожения сахаромицес карлсбергенсис (Saccharomyces carlsbergensis ) и дрожжи верхового брожения – сахаромицес церевизие (Saccharomyces cerevisiae ).
Первоначально были известны дрожжи верхового брожения, так как брожение проходило только при обычной температуре (как в виноделии, хлебопечении). желая получить напитки, насыщенные углекислым газом, стали проводить брожение при низких температурах. Под влиянием изменившихся внешних условий и были получены дрожжи низового брожения с определенными свойствами.
В пивоварении применяют разновидности дрожжей, отличающихся друг от друга одной или несколькими особенностями. Их получают из одной клетки. такие культуры называют расами (штаммами).
Дрожжи верхового брожения в процессе интенсивного броения всплывают на поверхность сбраживаемой жидкости, накапливаются в виде слоя пены и остаются в таком виде до конца брожения. Затем они оседают, образуя весьма рыхлый слой на дне бродильного аппарата. по своей структуре эти дрожжи относятся к пылевидным дрожжам, не слипающимися между собой в отличие от хлопьевидных низовых дрожжей, оболочки которых клейкие, что приводит к слипанию(агглютинация) и быстрому осаждению клеткок.
Дрожжи низового брожения не переходят в поверхностный слой пива- пену, а быстро оседают на дне бродильного аппарата.
Способность дрожжей к хлопьеобразованию имеет важное значение для технологии сбраживанияпивного сусла, так как способствует ускорению осветления пива и облегчает съём дрожжей из бродильного аппарата после брожения с последующим использованием их в качестве семенных дрожжей. Низкая температура при брожении содействует хлопьеобразованию.
Реакция среды сильно влияет на свойства дрожжей. Например, в кислой среде при рН менее 3 и в щелочной при рН более 8 хлопьевидные дрожжи становятся пылевидными. Хлопьевидные дрожжи по сравнению с пылевидными имеют более крупные клетки, меньше подвержены автолизу, дают большой прирост биомассы, обладают меньшей бродильной активностью, образуют меньше диацетила и высших спиртов в пиве, что положительно сказывается на его качестве.
Дрожжи низового брожения отличаются от дрожжей верхового брожения тем, что они полностью сбраживают раффинозу. Дрожжи низового брожения имеют оптимальную температуру для роста25 - 27С, минимальную 2-3С, а при 60-65С они отмирают. Максимальное развитие низовых дрожжей происходит при рН4,8-5,3. Кислород, растворенный в сусле,способствует размножению дрожжей,в то время как продукты брожения (этиловый спирт, диоксид углерода, высшие спирты, ацетальдегид, кислоты), а также повышенная концентрация сахара угнетают развитие дрожжей.
Пивные дрожжи должны отвечать следующим требованиям: Быстро сбраживать сусло, хорошо образовывать хлопья и осветлять пиво в ходе брожения, придавать пиву чистый вкус и приятный аромат.
Бродильную активность дрожжей определяют по степени сбраживания сусла. Степень сбраживания (V ) –это показатель, выраженный в процентах характеризующий отношение массы сброженного экстракта (Е-е) к массе сухих веществ в начальном сусле (Е):V = ((E -e )100)/ Е,Где е- содержание в пиве экстрактивных веществ, % к массе пива.
По степени сбраживания дрожжи делятся на сильно-, или высокосбраживающие (степень сбраживания 90-100%), среднесбраживающие (80-90%), слабо, или низкосбраживающие (менее 80%).
К сильносбраживающим относятся дрожжи рас: 11, F (получена в Чехии), штамм 8а(М). Дрожжи расы 11 нетребовательны к качаеству сырья, хорошо оседают, пиво характеризуется полным вкусом. Дрожжи расы F хорошо осветляют пиво, рпидают ему приятный аромат, устойчивы к инфекции. Дрожжи штамма 8а (М) имеют высокую бродильную активность, повышенный коэффициент размножения, хорошо оседают.Использование этих дрожжей дает возможность сократить длительность главного брожения с 7 до 5 сут и получить пиво с хорошим вкусом.
К среднесбраживающим относятся дрожжи рас 776, 41, 44, S (львовская), Р (Чехия),
А (рижская). Дрожжи расы 776 неприхотливы к сырью, их можно использовать для приготовления пива с применением несоложеных материалов. Готовое пиво имеет удовлетворительный вкус резкую хмелевую горечь. Дрожжи рас 41, 44, S , Робладают хорошей способностью оседания, вкус пива чистый мягкий, Дрожжи расы 44 дают возможность получать хорошее пиво при применении воды повышенной жесткости. Дрожжи рас F , A хорошо осветляют пиво, устойчивы к инфекции.
Для темных и специальных сортов пива применяют дрожжи верхового брожения.
Требования, предъявляемые к качеству дрожжей, не всегда удовлетворяются одной расой, поэтому в производстве применяют смесь рас или ведут брожение сусла отдельно на разных расах, а затем смешивают молодое пиво.
Разведение дрожжей чистой культуры .
Под разведением понимают увеличение массы дрожжей в количестве от массы в одной пробирке до массы маточных, необходимой для внесения в бродильный аппарат.
Весь процесс разведения состоит из двух стадий: лабораторной (разведение дрожжей в лаборатории) и цеховой (разведение дрожжей в отделении чистой культуры).
Лабораторная стадия состоит из нескольких последовательных пересевов. Вначале чистую культуру из пробирки пересевают в колбочки на стерильное сусло, затем проводят пересев дрожжей со стерильным сброженным суслом на новое стерильное сусло, объем которого от пересева к пересеву увеличивается в несколько раз. Лабораторная стадия заканчивается сбраживанием 6 л сусла в медной колбе Карлсберга в течении 5-6 сут при 7-8 о С.
Цеховая стадия- это разведение дрожжей на стерильном охмеленном сусле в специальных аппаратах.
На рисунке показана установка Грейнера для разведения чистой культуры дрожжей в цехе (трубопроводы не показаны). Установка состоит из стерилизатора 4 , двух бродильных цилиндров 3,число которых изменяется в зависимости от количества используемых дрожжей, резервуара для предварительного брожения 1 и посуда 2 для посевных дрожжей.
Стерилизатор и резервуар предварительного брожения оборудованы змеевиками для нагревания или охлаждения сусла, воздушными фильтрами и контрольно- измерительной аппаратурой.
Бродильные цилиндры имеют сосуды для посевных дрожжей вместимостью 10 л.
Стерилизатор предназначен для кипячения
сусла (стерилизация) и последующего его охлаждения, Бродильный цилиндр- для
первой стадии размножения, резервуар предварительного брожения- для
стерилизации и охлаждения сусла, а также проведения второй стадии размножения
чистой культуры. Температура воздуха в 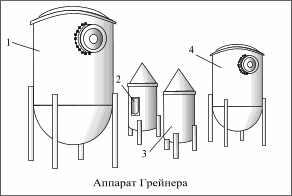 отделении
чистой культуры 8-9 о С.
отделении
чистой культуры 8-9 о С.
Разведение чистой культуры происходит следующим образом. В стерилизатор 4 из сусловарочного котла набирают горячее охмеленное сусло, кипятят его в течении 1 ч, затем охлаждают до8-12 о С. С помощью сжатого стерильного воздуха сусло подают в цилиндр 3, куда через специальный краниз медной колбы Карлсберга вводят чистую культуру, затем сбраживают в течении 3 сут.При этом дрожжи размножаются и увеличиваются в массе. к концу третьих суток резервуар предварительного брожения заполняют суслом, которое также нагревают до кипения, а затем охлаждают. Часть чистой культуры из бродильного цилиндра 3 отбирают на хранение в сосуд 2 для посевных дрожжей, где она хранится до до следующей разводки, а основную часть его перекачивают в резервуар 1,где осуществляют предварительное брожение при 9 о С в течение 3 сут.
В следующих циклах разведения дрожжи для посева в стерильное сусло, находящееся в бродильном цилиндре 3 берут из сосуда 2. Процесс разведения чистой культуры в установке Грейнера повторяют многократно до обнаружения в дрожжах посторонней микрофлоры.
Сбраживаемую массу из резервуара 1 перекачивают в специальный аппарат для предварительного брожения вместимостью 1000 дал, но наполненный на 1/3 суслом температурой 5-7 о С.Через 12 ч брожения в этот аппарат доливают еще 400 дал свежего охмеленного сусла и продолжают брожение еще 36 ч, поддерживая температуру 5-7С. Затем сбраживаемое сусло перекачивают в аппарат для главного брожения с 700 дал сусла, а через 1 сут заполняют его суслом до полной вместимости и ведут брожение обычным способом, контролируя температуру, концентрацию сусла и осветление. Осевшие при брожении дрожжи смывают, промывают холодной водой и используют в производстве как первую генерацию.
Дрожжерастительные аппараты перед началом работы стерилизуют паром в течение 45 мин под давлением 0,15- 0,17 МПа. Воздух, поступающий в стерилизатор, должен проходить через воздушные фильтры.
Спиртовое брожение — фундамент и начало всех напитков содержащих алкоголь, будь то вино, виски или пиво. Основа этого самого фундамента - сырье, вода и дрожжи. В этой статье мы расскажем о различных видах винных дрожжей, используемых в домашнем и промышленном виноделии. Рассмотрим, какие бывают дрожжи - дружественные, помогающие развивать богатство и разнообразие вин, и вражеские для винодела, угнетающие и портящие не только само вино, но и заражающие целые винодельни вместе с оборудованием.
Спиртовое брожение (оно же “ферментация”) - биохимический процесс, осуществляемый дрожжами, идеальным результатом которого является преобразование сахаридов (в основном, сахарозы, глюкозы и фруктозы) в этиловый спирт (основной продукт), углекислый газ и множество химических микроэлементов (нужных и не нужных, вредных и полезных побочных продуктов).
Дрожжи - микроскопические одноклеточные грибки. Cовременная микробиология разделяет их на более, чем полторы тысячи видов и еще тысячи подвидов, а уже они в свою очередь могут воспроизводить множество вариаций - в зависимости от результатов контролируемых и не контролируемых мутаций и перерождений (вы наверняка с этим сталкивались сами, если использовали одни и те же дрожжи по несколько раз, размножая их самостоятельно).
Человек еще с древних времен взял на вооружение спиртовое брожение, но пищевая промышленность и наука до сих пор открывают все новые и новые возможности и особенности использования дрожжей для производства этилового спирта. Множество усилий сконцентрировано именно в развитии рынка виноделия и микробиологии связанной с ним, это целая отрасль - энология. Энология занимается изучением и выведением бактерий, разработкой ферментов, исследованием и воспроизводством дрожжей обладающих нужными для виноделов качествами, позволяющие производить множество вин и винных напитков, открывая все новые грани и вкусы, а также сохраняя старые и редкие, ставшие историческим достоянием человечества.
Основные виды дрожжей (сахаромицетов - именно они друзья всех алковаров — виноделов, пивоваров и самогонщиков), используемых в производстве алкогольных напитков (в том числе в домашних условиях).
Таблица для наглядности. Здесь представлены некоторые расы дрожжей, вариации которых (разные штаммы одного и того же вида), популярны в промышленном (а некоторые и в домашнем) виноделии. Обратите внимание, что выбор конкретной расы обусловлен в том числе и условиями брожения. Некоторые рекомендуемые для виноделия расы винных дрожжей приведены в таблице (некоторые их иностранные аналоги доступны к покупке в нашем магазине).
«Saccharomyces cerevisiae» (Сахаромицес церевизия) — наиболее распространенный, разнообразный и "прирученный" вид дрожжей на данный момент в мире. Именно различные расы этого типа сахаромицета являются лидерами в сфере хлебопекарных, винных, пивных и спиртовых дрожжей. Они настолько разнообразны, а сферы их применения широки, что заслуживают отдельной статьи. К сожалению, не всегда в диком виноделии попадаются именно они, а учитывая что среди сахаромицетов церевизия существуют сотни подвидов негативно (в той или иной степени) влияющих на вино, то вероятность "удачного заражения" становится еще ниже, но не отсутствует;
«Saccharomyces vini» (Сахаромицес вини) - преимущественно они обитают на спелых (а особенно на поврежденных) ягодах винограда и в соках (незакрытых от внешнего воздействия). Нередко их можно обнаружить и в почве, в пищеварительной системе насекомых (особенно дрозофил, ос и пчёл), а также внутри винодельческого производства (в том числе и домашних виноделен) - на стенах, посуде и оборудовании. Однако, в домашнем виноделии они используются очень ограниченно, могут давать много нежелательных эффектов - например, помутнения вина и образования взвеси;
«Saccharomyces oviformis» (Сахаромицес овиформис) - чаще всего использование этой расы в виноделии может приносить пользу. Их используют для сбраживания сусла с высоким содержанием сахара, они хорошо подходят для производства сухих вин. Современные представители этой расы дрожжей имеют популярность в производстве шампанских вин.
Есть и отечественные расы: “Ленинградская”, “Киевская”. К недостаткам использования этих штаммов могут относиться - возобновление брожения в готовом вине (чаще всего полусладкого, но не только), а также помутнение и образование позднего осадка. Наиболее продуктивно использовать данные штаммы для производства Хереса - крепленого вина. Заточенные для этого представители (разновидность под названием “Oviformis Cheresiensis ”) - “Херес 96-К” и “Херес-20-С” - однако, они очень стремительно генерируют пленку на крепком вине (16-17% об.).
«Saccharomyces bayanus (uvarum)» (Сахаромицес баянус уварум) - чаще всего их можно встретить в плодовых винах и соках. Это очень неторопливые дрожжи - развиваются медленно, мутацию сложно контролировать из-за сложно-отличимых для современного уровня контроля микробиологических процессов в домашнем виноделии. Они не являются дрожжами с высокой степенью аттенюации (спиртообразования), однако имеют редкую особенность - повышенную устойчивость и сопротивление холоду. Что касается продуктов брожения - они почти идентичны тому, что набраживает вышеупомянутый штамм дрожжей S. Vini . Особенности - многие (но не все) расы этой разновидности способны образовывать плотнейший (не поддается взмучиванию) дрожжевой осадок, пена почти полностью отсутствует, дают повышенное содержание глицерина. Из наиболее популярных рас - “Новоцимлянская 3”, пока она почти недоступна для домашнего виноделия, но отлично себя зарекомендовала для производства полусладких вин.
Но далеко не все дрожжи одинаково полезны. Далее рассмотрим несколько опасных и подлых врагов виноделов - дрожжей, обладающих совсем недружелюбными свойствами.

«Pichia»/«Hansenula»/«Candida» и другие пленчатые — серьезные враги, вредители и виновники неудавшихся вин (особенно при использовании диких дрожжей). Главная их особенность - образование пленки на поверхности вина, особенно при аэробных условиях (гидрозатвор (водяной затвор) в помощь). Клетки этих зловредных дрожжей имеют нестабильную форму - встречаются эллиптические, овальные, колбасо- и булаво-видные формы и непропорционально удлиненные. Некоторые из них (Pichia и Hansenula) образуют споры, другие же - размножаются почкованием. Эти расы способны с высокой скоростью сбраживать винное сусло, окисляя его. Некоторые из них не могут набраживать достаточное для современного вина спирта, например, Hansenula - дает всего до 5% этанола.
В правильно подготовленном сусле они как правило не представляют опасности, т.к. содержатся в небольшом количестве. Если вы соблюдаете технологические условия подготовки и хранения виноматериалов (герметичность, стерильность материала и окружающей атмосферы), из которых будет сделано вино - опасаться нечего. Но вот при плохой подготовке (начинающие/незнающие виноделы любят не мыть виноград (не зная его микрофлору), использовать не подготовленную воду и тару) - штамм быстро начинает размножаться на поверхности. Это приводит к созданию уже на 2-3 сутки (иногда позже) глянцевой, а потом и складчатой (пупырчатой) пленки, которая может быть в широком цветовом диапазоне - от белого и мутно-прозрачного до серого цветов.
Если вино вдруг заразилось, то пленка это не самое худшее. Образование пленки означает новый этап сбраживания - сбраживание с окислением сахара. В процессе, обязательно образуются не только этиловые, а амиловые и бутиловые спирты, а также уксусные, масляные, янтарные кислоты и разнообразные эфирные соединения этих кислот. В результате чего, сусло получает характерный очень неприятный запах. Особенно велика вероятность, если вы используете мезгу - тогда для рас типа Pichia/Hansenula наступают наиболее благоприятные условия (поэтому мы рекомендуем для производства чачи использовать не винные, а более сильные фруктово-спиртовые дрожжи, которые способны нивелировать этот процесс).
Некоторые пленчатые дрожжи (часто встречающиеся в диком брожении - могут быть наоборот, очень спиртоустойчивыми (а еще и сульфито-устойчивыми), что поначалу нравится некоторым незадачливым виноделам. Они могут комфортно себя чувствовать даже при содержании спирта в 14% об. и 400-500 мг/л SO2. По этой же причине, в столовых винах (к которым есть доступ кислорода) они будут чувствовать себя очень комфортно, постепенно образовывая различные примеси которые приводят к порче вина (особенно домашнего и недопитого вина без консерванта). Они очень стремительно восстанавливают сульфаты и сульфиты, генерируя при этом большое содержание сероводорода и различных серных соединений, это является причиной неприятного тухлого запаха.
Цвель - одна из самых частых болезней вина дающих неприятный внешний вид (осадки, взвесь, островки, помутнение) и неприятный запах, зачастую появляется благодаря наличию в сусле активных дрожжей вышеупомянутых рас и пород. К тому же, они являются злейшими врагами производства сухих вин, шампанского и хереса - будьте осторожны!
Развитие и размножение злокачественных дрожжей в вине можно предотвратить ограничив доступ кислорода к нему (опять же, гидрозатвор) и установив правильные условия для хранения виноматериала - низкие температуры, а сами емкости должны быть заполнены полностью - не оставляя большого количества свободного пространства. Иногда может потребоваться своевременный долив - не стоит этим пренебрегать.
«Zygosaccharomyces» - еще один представитель зловредной (для винодела) микрофлоры. Эта раса дрожжей очень осмофильна - они прекрасно себя чувствуют в чрезвы чайно сладком сусле, развиваются даже при содержании сахара в 60 и даже 80 грамм на 100 мл. (вакуумное брожение сусла, различные домашние заготовки - мед, джемы, варенье). Они вызывают процесс ферментации в таких экстремальных условиях (представьте, какой это гидромодуль невероятный) и тем самым кардинально портят продукт. Что парадоксально, этот штамм не очень-то и спиртоустойчивый - образовывают в среднем не более 10-11% спирта, при этом бродят крайне долго, но за это время успевают полностью испортить некогда пригодный материал/продукт. Однако, современная наука нашла и им применение, правда ограниченное - “Расы Виерул, Май-копская, Краснодарская 40” - могут использоваться для снижения кислотности в очень кислом сусле, т.к. перерабатывают материал в основном путем брожения, а не окисления (как остальные зловредные).
«Saccharomycodes» - очередные вредители древнего и современного виноделия. Это сложные дрожжи, которые обладают крупной формой и неустойчивым типом размножения - могут и делиться и почковаться. Они обладаются спиртоустойчивостью до 12% (а по некоторым сообщениям и до 14%), а само сусло обогащают уксусно-этиловым эфиром, что губительно сказывается на выживаемости чистых культур дрожжей и могут вызывать различные побочные явления (например, внезапное возобновление брожения).
«Saccharomycodes ludwigii» - могут вызывать возобновление брожения даже высокосульфитированного сусла (не все консерванты являются панацеей). Также выдерживают очень высокое содержание SO2 в готовом вине и сусле.
«Hanseniaspora» (апикулятус) - чрезвычайно распространенные дрожжи, которые могут быть представлены как в виде спорогенов (Hanseniaspora apiculata) и аспорогенных грибов (Klocker apiculata). Если плод поврежден - очень вероятно, что они уже там, не брезгуют соками и крупными фруктами (да и мелкими тоже, просто чуть реже). Интересный факт: когда виноград созревает, они могут достигать до 99% всех дрожжей, которые там находятся (еще один повод в пользу чистоты - мойте виноград и весь виноматериал!). Сбраживающая способность низкая - всего около 4-7% об. спирта, однако очень много набраживают летучие соединения, уксусно-этиловый эфир, масляные, пропионовые и другие кислоты. Нередко являются причиной того, что сусло или вино (в особенности домашнее) недобродило. Они имеют взрывной рост и очень быстро размножаются в сусле, в несколько раз опережая рост других дрожжей - особенно благородных. Придают вину горечь, массу посторонних и при этом сильных запахов, а также уксусные оттенки. Делали домашнее шампанское или херес и попались “липнувшие" остатки? Это тоже их рук дело. Помочь может сульфитация (иногда повышенная) и длительное отстаивание.
«Torulopsis» - распространенная вредная для виноделия раса, особенно если речь идет о винограде. Они активно выделяются в уже забродившем виноградном соке, которое началось с диких дрожжей (т.н. благородной гнили). Их можно разделить на два основных вида (T. bacillaris и T. Candida). В последние годы встречалась информация о том, что они образуют споры, но в данный момент принято считать, что они размножаются почкованием. Они не часто появляются в вине, зато частый гость в виноградном соке. Способны сбраживать сусло до 12.5% оборотов этилового спирта. В плане биохимии они не так вредны и губительные как остальные дрожжи проявляющие себя при диком брожении, они образуют гораздо меньше зловредных химических элементов, но образуют - слизь. Согласитесь, мало кому будет приятно пить вино, в котором местами попадаются островки напоминающие кисель или что-то еще более склизкое. Они тоже являются осмофилами (им хорошо даже если содержание сахара 60-80 грамм на 100 мл) и любят высокую температуру, а повышение SO2 для них и вовсе не заметно.
«Rhodotorula» - это так называемые “розовые дрожжи”, их так называют из-за характерной окраски. Они не сбраживают сахара, а окисляют их, создавая при этом плотную розовую пленку. Способствуют окислению соков, образованию мути и выпадению в осадок десертных и полусладких (ну и сладких) вин. Интересная особенность - могут питаться парами спирта в воздухе, по этой причине часто оказываются “пойманными с поличным” на стенках виноделен и даже подвалов в виде розовой слизи.
Вместо вывода:
В произ-водственных условиях спонтанное брожение может вызвать нежелательные послед-ствия. Чтобы этого избежать и получить вино хорошего качества, брожение проводят на чистых культурах специально отселекционированных рас дрожжей, вводя их в сусло для направленного протекания процесса. В современном виноделии все же достаточно часто можно наткнуться на нейтральные и подходящие для виноделия дрожжи, они могут не быть истинными (идеальными, как чистые культуры дрожжей), но и не испортят вино. По этой причине нельзя однозначно считать, что спонтанное брожение обязательно приведет к порче вина, однако эта возможность существует всегда и порой эта вероятность может категорически возрастать. Например, если на одной кисти винограда оказываются зловредные киллер-дрожжи, при брожении они способны с огромной скоростью уничтожать чувствительных (1 клетка киллера может погубить в среднем 20 клеток благородных). К тому же, существует зловердно нейтральные (не участвующие во внутривидовой борьбе) расы, которые могут насытить вино лишними химическими соединениями, как правило плохо пахнущими.
Использование диких дрожжей часто чревато недобродом - внезапно сусло перестает "бурлить" или начинается "другое брожение" (пленчатое). Поэтому, если вы категорически используете дикие дрожжи - добавляйте диоксид серы , благородные штаммы как правило устойчивы к сульфитации, а вредные дрожжи - как правило нет (но, к сожалению, не все)
Вкусных всем вин!
Упоминающиеся товары в нашем .
ХЛЕБОПЕКАРНЫЕ ДРОЖЖИ - 2
Литература:
Семихатова Н.М. Хлебопекарные дрожжи: - М.: Изд-во «Пищевая промышленность», 1980 г. – 200 с. шифр 6П8.2 С30, инв. № 845314 хр
Матвеева И.В., Белявская И.Г. Биотехнологические основы приготовления хлеба. – М.: ДеЛи принт, 2001 г. – 150 с. (У Л.Ю.)
Влияние температуры .
Удельная скорость роста дрожжей при температуре:
20 °С = 0,149; 30 °С = 0,311; 36 °С = 0,342; 40 °С = 0,200; 43 °С = 0
Влияние активной кислотности среды .
Высокая скорость роста хлебопекарных дрожжей наблюдается при рН = 4,5 - 5,5. Подкисление среды во время выращивания хлебопекарных дрожжей до рН 3,0-3,5 и подщелачивание до 8,0 приостанавливает размножение дрожжевых клеток и ухудшает качество дрожжей.
Влияние химических веществ.
Рост дрожжей задерживается при содержании в среде более (%): сернистого ангидрида – 0,0025, фторида натрия – 0,002, нитритов – 0,0005, формалина – 0,001, карамелей – 0,1.
Задерживают рост дрожжей также кислоты при содержании их в среде более (%): щавелевая – 0,001, муравьиная – 0,0085, уксусная – 0,02, масляная – 0,005.
Также угнетают рост дрожжей соли вышеуказанных кислот при содержании их в среде более (%): 0,02-0,1. Концентрация солей кислот около 0,1 % тормозит рост дрожжей.
Действуют губительно соли металлов при содержании их в среде более (%): мышьяка – 0,0005, меди – 0,005, серебра – 0,000001. бактерицидное влияние солей металлов зависит от температурных условий, общей концентрации дрожжей, состава среды и её кислотности.
Скорость дрожжей также тормозят нитриты, образуемые бактериями, восстанавливающими нитраты в нитриты, которые являются ядом для дрожжей, в концентрации более 0.004 %.
Антибиотики не снижают активности дрожжей.
Влияние концентрации веществ во внешней среде при культивировании дрожжей .
5-6 % оптимальное содержание сахара в питательной среде при культивировании дрожжей.
Важное значение имеет осмочувствительность дрожжевых клеток, то есть их способность сбраживать сахара при повышенных концентрациях хлористого натрия (около 2 % к массе муки).
При выработке хлебобулочных изделий, в рецептуру которых входит сахар, важна устойчивость дрожжей по отношению к высокой концентрации сахаров (сахаротолерантность)
Влияние интенсивности аэрации и перемешивания на скорость роста дрожжей .
При выращивании дрожжей необходима аэрация питательной среды, которая количественно выражается в 0,8 г О 2 на 1 г углеродсодержащих питательных сред. Процесс расчёта интенсивности аэрации сложен и требуют отдельного изучения.
Ферменты дрожжей
Промышленное производство хлебопекарных дрожжей осуществляют, как правило, на мелассной массе, основной составной частью сахаров которой является сахароза. В связи с этим дрожжевая клетка активно индуцирует экзофермент ß- фруктофуранозидазу, легко выделяющийся в окружающую среду. Данный фермент всегда присутствует в клетке и сосредоточен с внешней стороны клеточной мембраны. В связи с этим гидролиз сахарозы происходит прежде, чем она проникает в дрожжевую клетку, активность фермента высока и проявляется с первых минут брожения полуфабрикатов.
Питательная смесь, в которой выращивают дрожжи, не содержит мальтозы, поэтому индукция фермента α-глюкозидазы (мальтазы) слаба. Эндофермент α-глюкозидазы локализуется в цитоплазме дрожжевой клетки. При сбраживании мальтозы углевод проникает внутрь клетки и там расщепляется на две молекулы глюкозы ферментом α-глюкозидазой.
Способность хлебопекарных дрожжей разрыхлять тесто зависит от активности зимазного комплекса клеток и от наличия сбраживаемых сахаров. Сахара в мучных полуфабрикатах хлебопекарного производства имеют несколько источников их происхождения – собственные сахара муки; сахара, получаемые под действием ферментов муки и дрожжей; сахара, добавляемые в полуфабрикаты по рецептуре.
Ввиду недостаточного количества собственных сахаров муки их технологическое значения невелико. В качестве источника углерода их достаточно только на начальный этап брожения полуфабрикатов. Источником сахара при созревании полуфабрикатов является крахмал, который под действием амилолитических ферментов муки расщепляется до α -β -декстринов и мальтозы. Основным «технологическим сахаром» в полуфабрикатах хлебопекарного производства, не содержащих в своем составе рецептурного сахара, является мальтоза.
Динамика сбраживания сахаров при использовании прессованных дрожжей в полуфабрикатах, рецептура которых не содержит сахарозы, представлена на рисунке.
Рис. Динамика сбраживания различных сахаров при брожении опары с применением прессованных дрожжей
При брожении опары одновременного сбраживания сахаров практически не происходит. В начале брожения дрожжевые клетки сбраживают глюкозу, а сбраживание фруктозы и мальтозы наступает через час и два часа соответственно. Зимазный комплекс
Зимазный комплекс ферментов дрожжей обеспечивает превращение моносахаров в спирт и диоксид углерода. Глюкоза сбраживается непосредственно, а фруктоза после изомеризации ее в глюкозу фруктоизомеразой дрожжей, которая является индуцируемым ферментом. Ферменты, сбраживающие глюкозу и сахарозу, являются конститутивными. Сахароза предварительно превращается в глюкозу и фруктозу под действием β -фруктофуранозидазы дрожжей, причем скорость ее инверсии очень высока.
При наличии мальтозы в среде дрожжевая клетка секретирует фермент мальтопермеазу, который осуществляет транспорт мальтозы внутрь клетки, и фермент α -глюкозидазу (мальтазу), расщепляющий мальтозу на две молекулы глюкозы, которая затем непосредственно сбраживается дрожжами при участии их зимазного комплекса с образованием диоксида углерода и этанола. Ферменты, участвующие в сбраживании мальтозы (мальтопермеаза и α -глюкозидаза), формируются только после того, как дрожжевые клетки оказываются в среде, содержащей этот дисахарид. Они являются индуцируемыми (адаптивными) ферментами.
Переключение дрожжей со сбраживания глюкозы на сбраживание фруктозы и мальтозы требует определенного периода, связанного с индукцией ферментов, поэтому скорость газообразования в полуфабрикатах в этот период незначительно снижается. После адаптации к сбраживанию мальтозы скорость газообразования в тесте опять возрастает до тех пор, пока не наступает недостаток мальтозы в среде (рис.).

Рис. Динамика скорости газообразования прессованных дрожжей в полуфабрикатах при опарном способе тестоприготовления
Фермент мальтопермеаза расположен в цитоплазматической мембране дрожжевой клетки, имеющей жидкостно-мозаичную структуру, является липидзависимым ферментом. Известно, что существует функциональная зависимость между активностью ферментных систем, расположенных в цитоплазматической мембране дрожжей, и ее микровязкостью. Таким образом, активность пермеазы, и следовательно, интенсивность ферментативных превращений внутри клетки зависит от микровязкости ее мембраны, воздействием на которую можно регулировать скорость биохимических процессов брожения.
Поскольку секреция индуцируемых ферментов дрожжей зависит от скапливающегося в среде субстрата (мальтоза), процесс адаптации клеток к мальтозной среде довольно продолжительный и это, вероятно, может отражаться на продолжительности сбраживания полуфабрикатов. Для ускорения процесса осахаривания крахмала муки в полуфабрикаты добавляют амилолитические ферментные препараты, что увеличивает содержание сбраживаемых сахаров в тесте и способствует интенсификации его созревания.
Высокой осахаривающей способности крахмала в мучных полуфабрикатах можно достичь путем изменения генетических свойств дрожжей, используемых в хлебопекарном производстве, а именно за счет регулирования биосинтеза и секреции определенных ферментов дрожжей.
Обобщенная модель спиртового брожения в пшеничных полуфабрикатах представлена на рис. 9.
Представленная схема, характеризующая роль дрожжей при производстве хлеба, свидетельствует о том, что эффективность полуфабрикатов зависит от целого комплекса биохимических превращений.

Гибридизация как эффективный метод отбора новых рас дрожжей
Гибридизация основана на способности дрожжей к половому размножению с образованием спор при неблагоприятных условиях – голодании, снижении температуры окружающей среды и т.д. В обычных условиях ряда производств дрожжи размножаются вегетативно - почкованием. Образующиеся в результате гибридизации гибриды обладают повышенной энергией размножения, чем исходные родительские расы.
Признаки, по которым отбирают гибридных сахаромицетов для производства:
Крупные клетки размером не менее 7х11 мкм;
Мальтазная активность не более мин;
Подъёмная сила не более 45 мин;
Устойчивость к мелассе 100 %
РАСЫ ДРОЖЖЕЙ
В настоящее время в дрожжевом производстве является общепринятым использование дрожжевых грибов вида Saccharomyces cerevisiae различных рас. Под расой понимают разновидность микроорганизмов, которые, сохраняя все основные признаки данного вида, отличаются второстепенными, но стойкими свойствами, характеризующими их производственные особенности. Очень часто расы называют штаммами, что неправильно, ибо штамм – это также разновидность данного вида, апробированная только в лабораторных условиях (Семихатова Н.М., 1980). Расой или штаммом называют отдельные разновидности микроорганизмов в пределах одного и того же вида, различающиеся между собой второстепенными признаками. При этом расы имеют стойкие второстепенные признаки, а штаммы нестойки и могут быть утрачены при росте на новой среде (Матвеева И.В., Белявская И.Г., 2001).
В связи с совершенствованием дрожжевого производства и процессов хлебопечения к дрожжам предъявляются ныне все новые и новые требования. Изменяются и взгляды на выбор признаков, характеризующих активную производственную расу. Прежде отбор осуществлялся в основном по культуральным и морфологическим признакам, а в настоящее время в основу его положены характеристики биохимических и ферментативных свойств дрожжей.
Производственные культуры дрожжей должны обладать высокой удельной скоростью роста, что особенно важно при многофазных технологических режимах приготовления хлеба, предусматривающих длительное приготовление полуфабрикатов, высокой активностью ферментов.
Характеристики морфологических и физико-химических свойств и технологических показателей отдельных штаммов дрожжей, применяемых в хлебопекарном производстве, приведены ниже.
Изначально в 1939 г выведена Раса Томская 7 Е.А. Плевако и Н.Г. Макаровой из прессованных дрожжей Томского дрожжевого завода. Эта раса характеризуется устойчивостью к составу мелассных сред, требовательностью к ростовым веществам, в частности к витаминам. Прессованные дрожжи, полученные на этой расе, стойкие при хранении, обладают высокой β -фруктофуранозидазной активностью, но слабой α -глюкозидазной активностью (мальтазная активность более 160 мин).
Раса Одесская 14 выделена в 1958 г. на Одесском дрожжевом заводе 3.И. Вишневской из образца импортных сушеных дрожжей. Культура отличается высокой генеративной активностью. Дрожжи устойчивы к высушиванию, в прессованном виде стойки при хранении. Мальтазная активность составляет 95 мин, зимазная – 45 мин. Культура требовательна к составу питательных сред, особенно к ростовым веществам. Однако, благодаря высокой урожайности и ферментативной активности, она нашла широкое распространение в промышленности.
Штамм Л-441 выведен в ЛО ГосНИИХП путем отбора на основе естественной изменчивости дрожжей расы Одесская 14. Штамм Л-441 характеризуется высокой продуктивностью, сбраживает раффинозу, устойчив к вредным примесям и патогенным микроорганизмам, имеет высокую удельную скорость роста и обеспечивает хорошие свойства товарных хлебопекарных дрожжей: подъемная сила 44-45 мин, мальтазная активность 92-95 мин, стойкость при температуре 35 °С свыше 96 ч.
Штамм Я-1 выведен на Янгиюльском дрожжевом заводе из производственной чистой культуры дрожжей расы 14 путем направленного отбора. Штамм испытан в производственных условиях в течение ряда лет. Культура обладает высокой генеративной активностью и устойчивостью к повышенной температуре выращивания (37-38 °С), что очень важно для заводов, находящихся в южных районах страны. Подъемная сила товарных дрожжей – 40-47 мин, зимазная активность 32-44 мин.
Раса Киевская 21 выделена в 1960 г. М.К. Рейдман из импортных сушеных дрожжей методом многократной активации с биогенными стимуляторами. Культура нетребовательна к ростовым веществам, хорошо переносит высушивание, обладает хорошей зимазной (60 мин) и мльтазной (100 мин) активностью.
Гибридные расы 176, 196-6 и 262 отвечают основным требованиям, предъявляем к производственным дрожжам, и рекомендованы для использования в промышленности: мальтазная активность 65-75 мин, зимазная 42-57 мин, высокая скорость роста.
Селекционированы новые штаммы 739, 743, 608, 616, 722 , отливающиеся высокой активностью ферментов. Выведен штамм ЛВ-7, используемый для производства прессованных и сушеных дрожжей. Штамм характеризуется повышенной устойчивостью к примесям мелассы и микрофлоре, инфицирующей дрожжевое производство, отличается повышенной продуктивностью и превосходит аналоги по концентрации трегалозы в 2 раза. Показатель подъемной силы прессованных дрожжей штамма ЛВ-7 составляет 43-47 мин, осмочувствительность – 6-10 мин .
Штамм дрожжей 616 используется для производства сушеных дрожжей и превосходит расу 14 по активности ферментных систем дрожжей. Мальтазная активность дрожжей составляет 67 мин, зимазная – 55 мин.
Штамм 722 отличается хорошей мальтазной (54 мин), зимазной (43 мин) активностью, подъемной силой (46 мин) и осмочувствительностью (5-10 мин) .
Штамм 739 характеризуется высокой продуктивностью, повышенной ферментной активностью. Дрожжи полностью сбраживают глюкозу, фруктозу, сахарозу, мальтозу, раффинозу, галактозу. Зимазная, мальтазная активности и подъемная сила дрожжей составляют соответственно 54, 61 и 56 мин .
Штамм дрожжей Saccharomyces cerevisiae 39/15 обладает хорошей бродильной активностью, его применение позволяет сократить продолжительность брожения теста на 35 мин.
Для производства сушеных дрожжей используется штамм Saccharomyces cerevisiae 93, обладающий высокой продуктивностью, активным комплексом ферментов. Зимазная активность составляет 45 мин, мальтазная – 53 мин, подъемная сила – 45 мин.
Гибридный штамм 512 получен скрещиванием расы XII и штамма Saccharomyces diastaticus , является триплоидом и характеризуется повышенным синтезом витаминов Д (эргостерина) – 2,8; В 1 – 34; В 2 – 20; В 6 46, РР – 36 (мкг/клетка). Показатели зимазной, мальтазной активности и осмочувствительности составляют 70, 200 и 14 мин соответственно.
Штамм 5 получен в результате скрещивания клеток штамма дрожжей «Яблочный-3», применяемого для сбраживания яблочного сока, и штамма 722, используемого в производстве сушеных хлебопекарных дрожжей. Отличительной особенностью штамма является высокая бродильная активность. Показатели зимазной, мальтазной активности и осмочувствительности составляют 85, 95 и 15 мин.
Штамм 69 получен в процессе скрещивания расы дрожжей «Джам-булская-60» и штамма 10, выделенного из сушеных дрожжей французского производства. Штамм 69 обладает высокой скорость роста, зимазной и мальтазной активностью соответственно 45 мин и 80 мин, а также устойчивость к повышенной температуре (40-45° С).
Представителем другого вида рода Saccharomyces являются дрожжи Saccharomyces minor , встречающиеся в ржаных заквасках. Это мелкие дрожжи круглой или слегка овальной формы, впервые выделенные и описанные в 1872 г. Энгелем. Они сбраживают и усваивают глюкозу, фруктозу, сахарозу, галактозу, раффинозу, не сбраживают и не усваивают лактозу, ксилозу, арабинозу, глицерин, манит, не расщепляют крахмал и клетчатку. Характерная особенность данного вида заключается в том, что он не сбраживает и не усваивает мальтозу и простые декстрины. Температурный оптимум для них составляет 25-28 °С и повышение температуры до 35 °С действует угнетающе. Дрожжи Saccharomyces minor отличаются большей кислотоустойчивостью (хорошо развиваются при кислотности 14-16° и рН 3,0-3,5) и спиртоустойчивостью, в отличие от Saccharomyces cerevisiae .
В настоящее время продолжаются работы по выведению новых штаммов дрожжей с применением современных методов: индуцируемого мутагенеза, гибридизации, адаптации. Это способствует эффективной селекции чистых культур микроорганизмов с закрепленными качественными признаками, необходимыми для реализаций современных технологий приготовления хлебобулочных изделий.
Виды хлебопекарных дрожжей
Для приготовления хлебобулочных изделий применяются хлебопекарные прессованные, сушеные, инстантные дрожжи, дрожжевое молоко, жидкие заквасочные дрожжи.
Прессованные дрожжи – это технически чистая культура дрожжей Saccharomyces cerevisuie , сформированная в брикеты влажностью 61-75 %. Культура выращивается на специальных питательных средах путем накопления биомассы маточных и засевных дрожжей в условиях интенсивной аэрации среды до получения товарных дрожжей прессованием или вакуумированием. В одном грамме прессованных дрожжей содержится 10-15 млрд. клеток.
Сушеные дрожжи – это высушенные до влажности 8-10 % при определенных условиях прессованные дрожжи, применяются после предварительной регидратации.
Быстрорастворимые (инстантные) дрожжи – высокоактивные сушеные дрожжи, не требующие регидратации перед внесением в тесто, приготовленные на основе определенных штаммов сахаромицетов с использованием современных условий культивирования, методов высушивания и защитных добавок и/или эмульгаторов.
Дрожжевое молоко (сепарированные дрожжи ) – дрожжевая суспензия концентрацией 400-450 г/л, полученная после сепарации и используемая взамен прессованных дрожжей.
Жидкие дрожжи – специально приготовленный на хлебозаводе полуфабрикат на основе осахаренной заварки, заквашенной термофильными молочнокислыми бактериями, с последующим выращиванием на ней дрожжей вида Saccharomyces . Жидкие дрожжи используются в качестве биологического разрыхлителя геста или как средство улучшения качества хлеба. В 1 мл жидких дрожжей содержится 70-120 млн. клеток.
Развитие современных технологий хлебопекарного производства требует использования дрожжей, адаптированных для использования к конкретным технологическим схемам, поэтому рядом предприятий и фирм выпускаются дрожжи осмотолерантные, полусухие замороженные, чувствительные к холоду, устойчивые к пропионату кальция, а также для использования в готовых смесях для хлебобулочных изделий.
Осмотолерантные дрожжи предназначены для приготовления теста с содержанием сахара-песка в рецептуре более 10 % к массе муки. Особенности осмотолерантных дрожжей заключаются в низком содержании инвертазы, способности синтезировать трегалозу и глицерин, что позволяет снизить осмотическое давление и компенсировать потери внутриклеточной воды.
Дрожжи полусухие замороженные предназначены для применения в технологии быстрозамороженных тестовых полуфабрикатов для булочных и сдобных изделий. Содержание сухих веществ в них составляет 75-77 %. В процессе производства дрожжей после сушки их замораживают, что придает им большую стабильность при хранении. Особенности полусухих замороженных дрожжей заключаются в замедленной интенсивности начала процесса брожения и в стабильности их свойств в замороженном тесте при низкотемпературном хранении.
Дрожжи, чувствительные к холоду , характеризуются чрезвычайно низкой ферментативной активностью в температурном диапазоне от 4 до 12 °С и стандартной активностью при температуре 30-40 °С. Это позволяет использовать их для приготовления теста, предназначенного для розничной торговли. Тестовые заготовки, приготовленные с этими дрожжами, могут храниться в течение нескольких дней при температуре 3-7 °С, не подвергаясь изменениям, сопутствующим процессу брожения, вследствие чего отпадает необходимость их быстрого замораживания.
Дрожжи, устойчивые к пропионату кальция , характеризуются повышенной кислототолераитностью и адаптивностью к тесту, приготовленному с добавлением пропионата кальция как средства предотвращения картофельной болезни хлебопекарных изделий.
Дрожжи, предназначенные для производства готовых смесей (премиксов ) , способны храниться при доступе кислорода и влаги, а также не требуют предварительной гидратации. Такими свойствами дрожжи обладают из-за особенного строения защитных гранул, имеющих специальную оболочку и характеризующихся высокой пористостью структуры, что способствует быстрому растворению гранул и полуфабрикатах хлебопекарного производства.
Дезактивированные дрожжи – продукт, не обладающий сбраживающей способностью, но имеющий ферментативную активность. Эти дрожжи являются натуральным улучшителем восстановительного действия для теста, которому необходимо придать эластичные свойства и увеличить растяжимость.
Эффективность применения различных видов дрожжей определяется знанием основных кинетических закономерностей сбраживания сахаров, воздействием параметров окружающей среды, особенностями метаболизма дрожжей в зависимости от состава питательной среды и обусловливается физиологическими, биологическими и технологическими свойствами дрожжей.
Прессованные дрожжи применяются при приготовлении пшеничного теста и теста из смеси ржаной и пшеничной муки в количестве от 0,1 до 8 % к массе муки в зависимости от рецептуры, способа производства и параметров технологического процесса.
Прессованные дрожжи вносятся в полуфабрикаты в виде предварительно приготовленной дрожжевой суспензии в воде при соотношениях от 1:2 до 1:4. Применение сушеных дрожжей включает предварительную стадию регидратации, иногда и активации. Для инстантных дрожжей не требуется предварительная подготовка, их вносят в тесто в сыпучем виде. Сравнительная характеристика при использовании различных видов хлебопекарных дрожжей приведена в табл. 1.
Важными факторами, которые находятся в зависимости от количества дрожжей в тесте и их активности, являются параметры технологического процесса – продолжительность и температура брожения полуфабрикатов. При сокращении процесса брожения теста количество дрожжей увеличивается. Отмечена прямая закономерность между величиной температурного коэффициента брожения и температурой брожения: при повышении температуры от 25 до 35 °С интенсивность брожения увеличивается примерно в 2 раза.
Дозировка дрожжей зависит от способа тестоприготовления, определяющим параметром которого является продолжительность процесса. В практике хлебопекарного производства в зависимости от способа приготовления теста используются следующие количества прессованных дрожжей: при опарном способе – 0,5-1,0 %; безопарном способе – 2,0-2,5 %; однофазных ускоренных способах – 3,0-6,0 % к массе муки.
Практическое значение имеет дифференциация сбраживающей активности дрожжей по отношению к различным сахарам в зависимости от способа тестоприготовления и, следовательно, продолжительности брожения полуфабрикатов. Для опарного и безопарного способов (общая продолжительность созревания которых составляет 210-350 мин) для достижения оптимальных свойств теста и хорошего качества хлеба имеет значение высокая мальтазная активность дрожжей. В процессе брожения опары, продолжительность брожения которой составляет 180-240 мин, происходит адаптация дрожжевых клеток к анаэробной мальтозно-мучной среде, поэтому интенсивность газообразования в тесте в значительно меньшей степени зависит от исходной мальтазной активности дрожжей по сравнению с безопарным способом.
При реализации ускоренных технологий, исключающих брожение теста в массе и имеющих общую продолжительность созревания полуфабрикатов 70-100 мин, индуцирование α -глюкозидазы дрожжами, которое начинается как правило через 70-90 мин от начало процесса брожения теста, не может оказать значительного воздействия на ход технологического процесса. Таким образом, приоритетное значение имеет высокая зимазная активность дрожжей. Рекомендацией при использовании ускоренной технологи приготовления хлебобулочных изделий является внесение в тесто не менее 2 % сахара-песка.
Важным фактором, влияющим на количество дрожжей в тесте, является рецептура, то есть количество сахара и жировых продуктов. Наличие в тесте сахара и жиросодержащих продуктов влияет на ферментативную активность дрожжей и, следовательно, на их количество. При внесении сахара-песка в количестве более 7 % к массе муки в тесте начинаются процессы плазмолиза дрожжевых клеток, вызывающего снижение их жизнедеятельности. Добавление в тесто жировых продуктов в количестве более 5 % вызывает снижение газообразования, обусловленное адсорбированием на поверхности дрожжевых клеток жира, что замедляет или останавливает прохождение растворимых питательных веществ через клеточную оболочку, нарушая процессы метаболизма дрожжей. Этим обусловлены рекомендации увеличения количества дрожжей в тесте для сдобных изделий до 4-6 % к массе муки или введения в технологический процесс стадии отсдобки теста, предусматривающей внесение сахара и жировых продуктов на последней стадии брожения теста.
Выбор вида и оптимальной дозировки дрожжей, продолжительности брожения полуфабрикатов хлебопекарного производства основывается на закономерностях, происходящих при их брожении, знании биотехнологических свойств различных видов дрожжей, механизмов влияния рецептурных компонентов во взаимосвязи с параметрами технологического процесса, способами тестоприготовления.
Расы пивных дрожжей
В пивоварении используют дрожжи низового брожения, приспособленные к сравнительно низким температурам. Пивные дрожжи должны быть микробиологиче-ски чистые, а также обладать способностью к хлопьеобразованию, быстро оседать на дно бродильного аппарата и давать прозрачный напиток с определенными вкусом и ароматом. К сильносбраживающим и легко дающим хлопья относятся пив-ные дрожжи низового брожения Фроберг (Saccharomyces cere-visiae Froberg), дрожжи рас V и 776.
На пивоваренных заводах большое распространение получили дрожжи расы 776, которая была выведена в начале XX в. Эти дрожжи считаются пригод-ными особенно для сбраживания сусла, приготовленного с добав-кой несоложеных материалов или из солода, полученного солодованием ячменей с невысокой степенью прорастаемости. Дрожжи расы 776 –среднесбраживающие, за период главного брожения на сусле концентрацией 11% образуют примерно 2,7% СО 2 . Клетки яйцевидной формы, длиной 8-10 мкм и ши-риной 5-6 мкм. Прирост дрожжевой массы 1: 5,4. Способность к осветлению удовлетворительная.
Из других дрожжей на пивоваренных заводах применяются расы 11, 41, 44, S-Львовская и др, различающиеся по бродиль-ной энергии, способности к осаждению и энергии роста.
Дрожжи расы 11 – сильносбраживающие, с хорошей способ-ностью к осветлению. Пиво, полученное с применением дрожжей расы 11, имеет хороший вкус. Эта раса получила широкое рас-пространение на пивоваренных заводах.
Дрожжи расы 41 – среднесбраживающие, с хорошей способ-ностью к осаждению. При сбраживании сусла расой 41 получа-ется мягкое пиво с чистым вкусом.
Дрожжи расы 44 – среднесбраживающие. Способность к осаждению хорошая. Сообщают пиву полноту вкуса и дают хорошие результаты при применении в производстве воды с по-вышенной жесткостью.
Дрожжи расы S – среднесбраживающие. Способность к осаждению хорошая. Дают пиво с мягким чистым вкусом.
Дрожжи расы Р – среднесбра-живающие, хорошо осветляют пиво и обусловливают приятный чистый вкус.
Дрожжи расы F характеризуются хо-рошей способностью к осветлению и сообщают пиву приятный аромат. Раса устойчива к действию посторонних микроорга-низмов.
Дрожжи расы А (выделены на рижском пивоваренном заво-де «Алдарис») сбраживают сусло за 7-8 суток, хорошо осветля-ют пиво и устойчивы к инфекции.
Путем разных способов селекции во ВНИИ пивобезалко-гольной промышленности получен ряд сильносбраживающих штаммов дрожжей (28, 48, 102), обладающих значительно боль-шей бродильной энергией, чем дрожжи исходной расы 11.
Пивные дрожжи верхового брожения находят широкое при-менение в Англии при приготовлении Портера. Они применяют-ся также для приготовления Берлинского светлого пива и дру-гих напитков. Для приготовления Бархатного пива применяют штамм 191 К, интенсивно сбраживающий моносахариды и маль-тозу, но не сбраживающий сахарозу, рафинозу и лактозу.
^ Расы винных дрожжей
В виноделии ценятся дрожжи, быст-ро размножающиеся, обладающие свойством подавлять другие виды дрожжей и микроорганизмы и придавать вину соответст-вующий букет. Дрожжи, применяемые в виноделии, относятся к своеобразному виду Saccharomyces ellipsoideus. Клетки их имеют продолговато-овальную форму. Дрожжи энергично сбра-живают глюкозу, фруктозу, сахарозу и мальтозу. В различных местностях и из различных молодых вин выделено несколько отличающихся одна от другой разновидностей или рас этого вида. В виноделии почти все производственные культуры дрож-жей – своего, местного происхождения. К их числу относятся расы Магарач 7, Массандра 3, Пино 14, Кахури и многие другие. Наряду с этими расами применяются и некоторые иностранные, например раса Штейнберг, выделенная в Германии в 1892 и 1893 гг., и раса Шампань-Аи.
Большая часть винных дрожжей относится к дрожжам низо-вого брожения.
Для приготовления белых столовых вин применяются расы Пино 14, Феодосия 1/19, Алиготе, Рислинг Анапский.
Раса Пино 14 имеет клетки яйцевидной формы, хорошо сбра-живает виноградное сусло сахаристостью 20 % с образованием 11,57%об спирта; оптимальная температура развития и бро-жения 18: -25°С. Эта раса является холодостойкой и кислото-стойкой; оптимальная величина рН 2,9-3,9.
Раса Феодосия 1/19 – крупноклеточная, пылевидная, очень энергичная, быстро сбраживает виноградное сусло и хорошо дображивает его; имеет широкий температурный диапазон бро-жения (от 9 до 35°С) и может применяться как холодостойкая и как термостойкая.
Дрожжей Алиготе имеется несколько рас, и все они сильные, с высокой энергией брожения. К энергично сбраживающим от-носятся и дрожжи Рислинг Анапский.
Для приготовления крепких вин применяется раса Массанд-ра 3 с яйцевидной формой клеток, пылевидная; оптимальная величина рН 3,7-4,05; оптимальная температура брожения 18-20°С. Виноградное сусло с содержанием сахара 20% сбраживается полностью; при сбраживании концентрированного виноградного сусла (30% сахара) образует 11,8%об спирта и оставляет несброженным 8,7% сахара.
Раса Магарач 125, названная в ознаменование 125-летнего юбилея первых посадок винограда в институте «Магарач», при-меняется для получения крепких и десертных вин. Эта раса хо-рошо сбраживает высококонцентрированные виноградные сусла с содержанием сахара 27-30%, холодостойкая.
Раса Кахури 2 широко применяется для приготовления шам-панских виноматериалов и вин. Виноградное сусло с содержанием сахара 20% она сбраживает с образованием 11,4%об спирта, оста-ется несброженным 0,28% сахара. Эта раса довольно холодо-стойкая (при температуре 14-15°С сусло забраживает на 2-й день) и сбраживает хорошо; оптимальная величина рН 3,4-3,6.
Раса Шампанская 7, применяемая для шампанизации вина в бутылках, выделена из расы Кахури 5 и характеризуется об-разованием трудно взмучивающегося осадка; интенсивно сбра-живает при температуре 4-9°С, хотя сусло и забраживает только на 5-6-й день.
Из винных дрожжей наиболее холодостойкой считается раса Ленинградская, а наиболее термостойкой – раса Ашхабадская 3.
В производстве хереса применяются специальные расы дрож-жей, которые являются разновидностью вида Saccharomyces oviformis. Хересные дрожжи образуют на поверхности вина в неполных бочках пленку, благодаря развитию которой вино приобретает особые букет и вкус.
Путем тщательного отбора по наиболее важным производст-венным признакам выделено несколько рас хересных дрожжей (13, 15 и 20) с высокой пленкообразующей способностью. В дальнейшем из производства, применявшего расу Херес 20, была отселекционирована более эффективная раса Херес 20-С, которая нашла широкое применение на многих заводах по про-изводству хереса.
В плодово-ягодном виноделии применяются селекционирован-ные расы дрожжей, выделенные из различных плодово-ягодных соков. Плодово-ягодные соки богаты дрожжами, обладающими всеми необходимыми для производства качествами и биологи-чески приспособленными к условиям развития в исходных пло-дово-ягодных соках. Поэтому штаммы дрожжей, выделенных из земляничных соков, применяются для сбраживания земляничных соков, а штаммы дрожжей, выделенных из вишневых соков, при-меняются для сбраживания вишневых соков и т. д.
В плодово-ягодном виноделии получили распространение следующие штаммы: яблочные 46, 58, клюквенный 17, смородиновый 16, брусничные 3, 7, 10, малино-вые 7/5, 25, 28, 28/10, вишневые 3, 6, земляничные 7, 4, 9.
Назван-ные штаммы дрожжей обеспечивают нормальный ход брожения, полноту сбраживания, быструю осветляемость и хорошие вкусо-вые качества вина; они сбраживают глюкозу, фруктозу, сахаро-зу, мальтозу, галактозу и не сбраживают лактозу и маннит.
Успешно применяются в плодово-ягодном виноделии расы дрожжей Москва 30, Яблочная 7, Вишневая 33, Черносмородиновая 7, Малиновая 10 и Сливовая 21. Чистая культура дрожжей Москва 30 рекомендуется для сбраживания клюквенного сусла; Яблочная 7 и Вишневая 33 – для сбраживания яблочного сус-ла; Черносмородиновая 7 и Вишневая 33 – для сбраживания черносмородинового и вишневого сусла.
^
4 Химизм спиртового брожения. Вторичные и побочные продукты спиртового брожения
Спиртовое брожение представляет собой цепь ферментатив-ных процессов, конечным результатом которых является распад гексозы с образованием спирта и СО 2 и доставка дрожжевой клетке той энергии, которая необходима для образования новых веществ, используемых для процессов жизнедеятельности, в том числе для роста и размножения. По химическому характеру спир-товое брожение – каталитический процесс, происходящий под действием биологических катализаторов – ферментов.
Современная теория спиртового брожения является резуль-татом работ многих ученых различных стран мира.
Для выяснения процессов брожения большое значение имели работы выдающихся отечественных ученых: Лебедева, Костычева, Фаворского, Иванова, Энгельгардта.
По современным представлениям, спиртвое брожение – это сложный непрерывный процесс распада сахара, катализируемый разными ферментами с образованием 12 промежуточных продуктов.
1 Начальной стадией превращения глюкозы является ре-акция фосфорилирования ее при участии фермента глюкозиназы. К молекуле глюкозы присоединяется фосфатный остаток от молекулы АТФ, которая находится в клетках дрож-жей, и образуется глюкозо-6-фосфат, а АТФ превращается в АДФ:
С 6 Н 12 О 6 + АТФ → СН 2 О(Н 2 РО 3)(СНОН) 4 СНО+АДФ
Глюкоза Глюкозо-6-фосфат
В результате присоединения фосфатного остатка от молеку-лы АТФ к глюкозе реакционная способность последней возра-стает.
2 Глюкозо-6-фосфат путем изомеризации под действием фермента глюкозофосфатизомеразы переходит обрати-мо в форму фруктозы:
СН 2 О(Н 2 РО 3)(СНОН) 4 СНО → СН 2 О(Н 2 РО 3)(СНОН) 3 СОСН 2 ОН
Глюкозо-6-фосфат Фруктозо-6-фосфат
3 Далее под действием фермента фосфофруктокиназы со второй молекулы АТФ переносится еще один фос-форный остаток на фруктозо-6-фосфат и образуется фруктозо-1,6-дифосфат и новая молекула АДФ:
СН 2 О(Н 2 РО 3)(СНОН)3СОСН 2 ОН + АТФ →
Фруктозо-6-фосфат
→ СН 2 О(Н 2 РО 3)(СНОН) 3 СОСН 2 О(Н 2 РО) + АДФ
Фруктозо-1,6-дифосфат
Эфиры глюкозо-6-фосфат и фруктозо-6-фосфат образуют равновесную смесь, получившую название эфира Эмдена и со-стоящую на 70-75% из эфира Робисона (глюкозы) и на 25% из эфира Нейберга (фруктозы).
Образованием фруктозо-1,6-дифосфата заканчивается подготовительная стадия спиртового брожения с переносом макроэргических фосфатных связей и с преобразованием гексозы в лабильную оксиформу, легко подвергающуюся дальнейшим фер-ментативным превращениям.
4 Следующим важнейшим этапом является десмолиз – разрыв углеродной цепи фруктозодифосфата с образованием двух
молекул фосфотриоз. Симметричное расположение остатков фосфорной кислоты по концам молекулы фруктозы облегчает разрыв ее углеродной цепи как раз в середине. Фруктозодифосфат распадается при этом на две триозы: фосфоглицериновый альдегид и фосфодиоксиацетон. Реакция катализируется фер-ментом альдолазой и обратима:
СН 2 О(Н 2 РО 3)(СНОН) 3 СОСН 2 О(Н 2 РО) → CH 2 О(Н 2 Р0 3)СОСН 2 ОН +
Фруктозо-1,6-дифосфат Фосфодиоксиацетон
СН 2 0 (Н 2 РОз) СНОНСНО (4)
З-фосфоглицериновый альдегид
Главная роль в дальнейших превращениях при спиртовом брожении принадлежит 3-фосфоглицериновому альдегиду, но в сбраживаемой жидкости он обнаруживается лишь в незначи-тельном количестве. Это объясняется взаимным переходом ке-тозного изомера в альдозный и обратно под действием фермента триозофосфатизомеразы (5.3.1.1)
СН 2 0 (Н 2 Р0 3) СОСН 2 ОН;£ СН 2 0 (Н 2 Р0 3) СНОНСНО
Фосфодиоксиацетон З-фосфоглицериновый альдегид
По мере дальнейшего превращения фосфоглицеринового аль-дегида новые количества его образуются в процессе изомериза-ции фосфодиоксиацетона.
5. Последующим этапом является окисление двух молекул З^фосфоглицеринового альдегида. Эта реакция катализируется триозофосфатдегидрогеназой (1.2.1.12), коферментом которой яв-ляется НАД (никотинамид-аденин-динуклеотид). В окислении участвует фосфорная кислота среды. Реакция протекает по сле-дующему уравнению: 2СН 2 0 (Н 2 Р0 3) СНОНСНО + 2Н 3 Р0 4 + 2НАД Триозофосфатдегидрогеназа ->
З-фосфоглицериновый альдегид
->- 2СН 2 0 (Н 2 Р0 3) СНОНСОО w (H 2 P0 3) + 2НАД Н 2 (5)
1,3-дифосфоглицериновая йислота
Молекула 3-фосфоглицеринового альдегида присоединяет фосфат, а водород переносится на кофермент НАД, который восстанавливается. Энергия, освобождающаяся в результате окисления 3-фосфоглицеринового альдегида, аккумулируется в макроэргической связи образующейся 1,3-дифосфоглицериновой
Кислоты.
6. Далее фосфатный остаток 1,3-дифосфоглицериновой кисло-
ты, содержащий макроэргическую связь, при участии фермента
фосфоглицераткиназы (2.7.2.3) переносится на молекулу АДФ.
Образуется 3-фосфоглицериновая кислота, а АДФ, приобретая
дополнительную макроэргическую связь, превращается в АТФ:
2СН 2 0 (Н 2 Р0 3) СНОНСОО со (Н 2 Р0 3) + 2АДФ->2СН 2 0 (Н 2 Р0 3) CHOHCOOH+
1,3-дифосфоглицериновая кислота 3-фосфоглицериновая кислота
7. Затем под действием фермента фосфоглицеромутазы
(2.7.5.3) остаток фосфорной кислоты перемещается от третьего
углерода ко второму, и в результате 3-фосфоглицериновая кис-
лота превращается в 2-фосфоглицериновую кислоту:
2СН 2 (Н 2 Р0 3) CHOHCOOH ^t 2CH 2 0HCH0 (Н 2 Р0 3) СООН. (7)
3-фосфоглицериновая кислота 2-фосфоглицериновая кислота
8. Следующей стадией является дефосфорилирование 2-фос-
фоглицериновой кислоты. При этом 2-фосфоглицериновая кис-
лота под действием фермента энолазы (4.2.1.11) путем дегидра-
тирования (потери воды) превращается в фосфоэнолпировино-
градную кислоту:
2СН 2 ОНСНО (Н 2 Р0 3) СООН qt 2СН 3: СО со (Н 2 Р0 3) СООН + 2Н 2 0. (8)
2-фосфоглицериновая кислота Сосфоэнолпировиноградная кислота
При этом превращении происходит перераспределение внут-римолекулярной энергии и большая ее часть аккумулируется в макроэргической фосфатной связи.
9. Весьма нестойкая фосфоэнолпировиноградная кислота
легко дефосфорилируется, при этом остаток фосфорной кислоты
под действием фермента пируваткиназы (2.7.1.40) передается
вместе с макроэргической связью молекуле АДФ. В результате
образуется более устойчивая кетоформа пировиноградной кисло-
ты, а АДФ превращается в АТФ:
2СН 2: СО сю (Н 2 Р0 3) СООН + 2АДФ -* 2СН 3 ^ СОСООН + 2АТФ. (3)
Фосфоэнолпировиноградная Пировиноградная
Кислота кислота
10. Пировиноградная кислота под действием фермента пи-
руватдекарбоксилазы (4.1.1.1) декарбоксилируется с отщепле-
нием С0 2 и образованием уксусного альдегида:
2CH 3 ^ COCOOH -*2С0 2 + 2СН 3 СНО. (10)
Пировиноградная Уксусный альдегид
11. Уксусный альдегид при участии фермента алкогольдегид-
рогеназы (1.1.1.1) взаимодействует с НАД-Н 2 , образовавшимся
ранее при окислении фосфоглицеринового альдегида в фосфо-
глицериновую кислоту [см. уравнение (5)]. В результате уксус-
ный альдегид восстанавливается в этиловый спирт, а кофермент
НАД-Н 2 вновь регенерируется (окисляется в НАД):
2СН 3 СНО + 2НАД Н 2 Z 2СН 3 СН 2 ОН + 2НАД. (11)
Итак, завершающим этапом брожения является реакция восстановления уксусного альдегида в этиловый спирт.
Из рассмотренного цикла реакций спиртового брожения вид-но, что из каждой молекулы глюкозы образуется 2 молекулы спирта и 2 молекулы С0 2 .
В процессе спиртового брожения образуется четыре молеку-лы АТФ [см. уравнения (6) и (9)], но две из них затрачиваются на фосфорилирование гексоз [см. уравнения (1) и (3)]. Таким образом, запасается всего 2 г-моль АТФ.
Ранее указывалось, что на образование каждой грамм-моле-кулы АТФ из АДФ затрачивается 41,9 кДж, а в энергию двух молекул АТФ переходит соответственно 83,8 кДж. Следователь-но, при сбраживании 1 г-моль глюкозы дрожжи получают энер-гии около 84 кДж. В этом и заключается биологический смысл брожения. При полном расщеплении глюкозы на С0 2 и воду выделяет-ся 2874 кДж, а при окислении 1 г-моль глюкозы до С0 2 и Н 2 0 в процессе аэробного дыхания аккумулируется 2508 кДж, так как образующийся этиловый спирт еще сохраняет в себе потен-циальную энергию. Таким образом, с энергетической точки зре-ния брожение - процесс малоэкономичный.
Сбраживание отдельных Сахаров происходит в определенной последовательности, обусловленной скоростью их диффузии в дрожжевую клетку. Быстрее всех сбраживаются дрожжами глюкоза и фруктоза. Однако сахароза как таковая исчезает в сусле (инвертируется) еще в начале брожения. Она гидролизу-ется р-фруктофуранозидазой (3.2.1.26) оболочки дрожжевых клеток с образованием гексоз (глюкозы и фруктозы), которые легко используются клеткой. Когда в сусле почти не остается фруктозы и глюкозы, дрожжи начинают потреблять мальтозу.
§ 5. ВТОРИЧНЫЕ И ПОБОЧНЫЕ ПРОДУКТЫ СПИРТОВОГО БРОЖЕНИЯ
Все вещества, получающиеся в результате сбраживания са-хара дрожжами, за исключением спирта и С0 2 , относятся к вто-ричным продуктам спиртового брожения. Кроме них, имеются побочные продукты спиртового брожения, которые образуются не из сахара, а из других веществ, находящихся в сбраживае-мом субстрате. К ним относятся амиловый, изоамиловый, изо-бутиловый и другие спирты, известные под названием сивушного масла.
Из вторичных продуктов спиртового брожения известны глицерин, уксусный альдегид, пировиноградная, уксусная, ян-тарная, лимонная и молочная кислоты, ацетоин (ацетилметил-карбинол), 2,3-бутиленгликоль и диацетил. В аэробных условиях пировиноградная кислота является также исходным веществом для цикла трикарбоновых кислот (цикла Кребса), по которому из нее образуются уксусная, лимонная, яблочная и янтарная кислоты. Высшие спирты образуются тоже из пировиноградной кислоты путем аминирования ее до аланина, который в свою очередь переаминируется в соответствующую кетокислоту. В ус-ловиях спиртового брожения кетокислоты, восстанавливаясь, образуют высшие спирты. Поэтому вторичные и побочные про-дукты спиртового брожения строго разграничить нельзя.
Уксусный альдегид может испытывать дисмутацию с обра-зованием уксусной кислоты и этилового спирта (реакция Кан-ниццаро):
СН 3 СОН + СН 3 СОН + Н 2 0 = СНзСООН + СН 3 СН 2 ОН.
Одна из молекул альдегида окисляется в кислоту, а другая восстанавливается в спирт. В щелочной среде одна молекула
Уксусного альдегида вступает в окислительно-восстановитель-ную реакцию со второй молекулой уксусного альдегида; при этом образуются этиловый спирт, уксусная кислота и одновре-менно с ними глицерин, что выражается таким суммарным урав-нением:
2C 6 Hi 2 0 6 + Н 2 0 = 2СН 2 ОНСНОНСН 2 ОН + СН 3 СН 2 ОН + СН 3 СООН + 2С0 2 .
Глицерин образуется в небольшом количестве при спиртовом брожении. При изменении условий брожения его производство можно осуществить в промышленном масштабе.
Глицерин и уксусный альдегид являются промежуточными продуктами спиртового брожения. На последнем этапе нормаль-но протекающего процесса брожения происходит восстановле-ние значительной части уксусного альдегида в этанол. Но если уксусный альдегид связать сульфитом натрия, то направление спиртового брожения изменится в сторону образования больших количеств глицерина.
Удаление уксусного альдегида из сбраживаемой среды суль-фитом натрия представляется в следующем виде:
СН 3 СНО + Na 2 S0 3 + H 2 OW CH 3 CHONaHS0 2 + NaOH.
Уксусный альдегид, образовавшийся при декарбоксилирова-нии пировиноградной кислоты, в результате связывания суль-фитом не может служить акцептором водорода. Место уксусно-го альдегида занимает фосфодиоксиацетон, который получает водород от восстановленного НАД-Н 2 , образуя а-глицерофос-фат. Эта реакция катализируется ферментом глицерофосфатде-гидрогеназой. Под действием фосфатазы «-глицерофосфат де-фэсфорилируется, превращаясь в глицерин. Таким образом, в присутствии Na 2 S03 протекает глицерино-альдегидное броже-ние:
С 6 Н 12 0 6 = CH3CHO + СН 2 ОНСНОНСН 2 ОН + С0 2 .
Сахар Ацетальдегид Глицерин
С увеличением количества сульфита натрия, вводимого в сбраживаемую среду, соответственно увеличивается количество связанного альдегида и ослабляется образование этанола и С0 2 .
Образование кислот и ацетоина. Янтарная кислота образует-ся дегидрированием и конденсацией двух молекул уксусной кис-лоты с одной молекулой уксусного альдегида (гипотеза В. 3. Гва-ладзе и Женавуа):
2СН 3 С00Н + СН 3 СНО -* С00НСН 2 СН 2 С00Н + СН 3 СН 2 ОН.
В процессе спиртового брожения янтарная кислота образует-ся также дезаминированием глютаминовой кислоты. Акцепто-ром водорода в этой реакции является триозоглицериновый аль- Дегид, поэтому реакция дезаминированйя сопровождается одно-временным накоплением глицерина:
C 6 Hi 2 0 6 + COOHCH2CH2CHNH2COOH + 2Н 2 0 = СО0НСН 2 СН 2 С0ОН -Ь
Глюкоза Глютаминовая кислота Янтарная кислота
2СН 2 ОНСНОНСН 2 ОН 3 + NH 3 + С0 2 .
Глицерин
Амми>ак потребляется дрожжами на синтез белка, а глице-рин и янтарная кислота при этом выделяются в среду.
Образование лимонной кислоты, по Лафону, происходит из. девяти молекул уксусного альдегида:
9СН 3 СОН + 4Н 2 0 = (СН 2 СООН) 2 С (ОН) СООН + 6СН 3 СН 2 ОН.
Лимонная кислота
Образование молочной кислоты объясняют восстановлением пировиноградной кислоты:
СНзСОСООН + Н 2 -> СН 3 СН (ОН) СООН.
Пировиноградная Молочная кислота
Однако полагают более вероятным ее образование в резуль-тате гидролиза промежуточного продукта спиртового броже-ния - фосфоглицеринового альдегида:
СНОСНОНСН 2 ОР0 3 Н 2 + Н 2 0 -* СН 3 СН (ОН) СООН + Н 3 Р0 4 .
Фосфоглицериновьш Молочная кислота
Альдегид
Конденсацией уксусной кислоты с ацетальдегидом объясня-ют образование ацетоина:
1) СНзСООН + СН 3 СНО->-СНзСОСОСНз + Н 2 0;
Диацетил
2) СН3СОСОСН3 + СНзСНО -4 СН3СОСНОНСН3 + СНзСООН.
Сначала образуется диацетил; затем путем дисмутации со-пряженного окисления-восстановления за счет воды диацетила с ацетальдегидом образуется ацетоин.
При восстановлении ацетоина образуется 2,3-бутиленгли-коль:
СН 3 СОСНОНСНз + НАД ■ Н 2 *± СН 3 СНОНСНОНСН 3 + НАД.
Механизм образования некоторых вторичных продуктов спиртового брожения еще не совсем ясен, однако не подлежит сомнению, что уксусный альдегид является основным исходным веществом для синтеза вторичных продуктов брожения.
Среди вторичных продуктов преобладают уксусная и янтар-ная кислоты, а также 2,3-бутиленгликоль и уксусный альдегид-В весьма малых количествах находятся ацетоин и лимонная кис-лота.
^ Образование высших спиртов. Своеобразным побочным про-дуктом спиртового брожения являются высшие спирты. Иссле-дованиями И. Я- Веселова установлено, что высшие спирты при брожении возникают главным образом в период размножения
Дрожжей. В этот период интенсивность обмена веществ связана с образованием кетокислот из продуктов превращения углево-дов с переаминированием их. Переаминирование заключается в обмене радикалами CH(NH) 2 и СО между аминокислотой и ке-токислотой. Так, образование аланина из лейцина и пировино-градной кислоты представляется в таком виде:
(CH 3)2CHCH 2 CHNH 2 COOH -f CH3COCOOH -> СН 3 СНСН 3 СН 2 СОСООН +
Лейцин Пировиноград- Изонронилвиноградная
Ная кислота кислота
CH 3 CHNH 2 COOH.
Изопропилвиноградная кислота, подвергаясь (аналогично
Пировиноградной кислоте в схеме спиртового брожения) декар-
Боксилированию, превращается в изовалериановый альдегид,
Который восстанавливается до изоамилового спирта:
СН 3 СНСН 3 СН 2 СОСООН -> (СН 3) 2 СНСН 2 СНО -*- СН 3 СНСН 3 СН 2 СОН.
Изопропилвиноградная Изовалериановый Изоамиловый
Кислота альдегид спирт
Аналогичным путем из изолейцина образуется амиловый, из валина изобутиловый спирты.
Таким образом, синтез новых аминокислот происходит с уча-стием пировиноградной кислоты, которая играет роль главного моста между углеводным и азотистым обменом в дрожжевой клетке.
На образование высших спиртов влияет ряд факторов. По мере увеличения или уменьшения нормальной температуры бро-жения количество высших спиртов сокращается. При примене-нии рН сбраживаемой среды от 3 до 5 накопление высших спиртов увеличивается, а при дальнейшем увеличении рН - уменьшается. Аэрация среды благоприятствует синтезу высших спиртов: в аэрируемой среде содержание изобутилового и изо-амилового спиртов увеличивается. Введение аминокислот в сбраживаемую среду, содержащую сахарозу, также стимулиру-ет накопление высших спиртов. Образование сивушного масла в культуральной среде возрастает с накоплением биомассы дрожжей. Содержание высших спиртов в сбраживаемой среде можно уменьшить путем торможения размножения дрожжей.
^ Образование эфиров. При участии эстераз дрожжей протека-ют реакции этерификации, в которых участвуют спирт и кисло-ты. В общем виде реакция этерификации представляется так: RCH 2 OH + RiCOOH -> RCOOCH 2 R! + Н 2 0.
Так, при взаимодействии этанола с уксусной кислотой обра-зуется уксусноэтиловый эфир (этилацетат):
С 2 Н 5 ОН + СН3СООН 5s CH 3 C0 2 C 2 H s + Н 2 0.
Образование эфиров протекает легче, когда компонен тами этой реакции являются альдегиды. Альдегиды легко подверга- ются окислительно-восстановительным превращениям и дают на-чало образованию кислот, спиртов, эфиров. При этом все пре-вращения альдегидов могут осуществляться как самостоятель-ные реакции без затраты энергии.
Так, образование эфиров может происходить за счет альде-гидов:
RCHO + HOCRi ->- RCOOCHjRj.
Альдегиды могут претерпевать альдольную конденсацию: СН 3 СНО + С"НзСНО = СН 3 СНОНСН 2 СНО.
Образующееся вещество содержит одновременно альдегид-ную и гидроксильную (алкогольную) группы.
При взаимодействии со спиртом уксусный альдегид превра-щается в диэтилацеталь:
СН 3 СНО + 2С 2 Н 5 ОН -* СНзСН (ОС,Н 5) 2 + Н 2 0.
В результате сбраживания Сахаров и всех сопутствующих, процессов сусло в пивоварении и виноделии превращается в го-товый продукт (пиво, вино). Все находящиеся в нем вещества обусловливают его аромат и вкус. Так, высшие спирты (пропи-ловый, амиловый, изоамиловый, тирозол, триптофол) обладают характерным запахом и дают сложные эфиры, которые уже имеют более приятные, смягченные запахи. 2,3-Бутиленгликоль. и глицерин обладают сладким вкусом.
В спиртовом производстве сброженную среду называют зре-лой бражкой, из которой путем отгонки на брагоректификацион-ных аппаратах получают спирт. Образующиеся в ходе брожения этиловый спирт и СОг выходят из клеток наружу в сбраживае-мую среду. Спирт хорошо растворяется в сбраживаемом сусле, в любых соотношениях и равномерно распределяется в нем. СОг! же сначала растворяется в сусле, а по мере насыщения его вы-деляется в виде газовых пузырьков. На поверхности газовых пузырьков появляется адсорбционный слой из поверхностно-ак-тивных веществ (белки, пектин). При слипании отдельных пу-зырьков получаются ячейки пены. Постепенно поверхность-сбраживаемого сусла покрывается пеной.д
Для сбраживания пивного сусла употребляются дрожжи верхового и низового брожения, размножающиеся почкованием. Эти дрожжи относятся к семейству сумчатых грибков - эндомицетов и входят в группу сахаромицетов.
Сбраживание сусла дрожжами верхового брожения имеет весьма характерные внешние признаки. Верховые дрожжи во время брожения поднимаются кверху и образуют шапку пены. К концу брожения часть дрожжей садится на дно чана.
Бродильная способность верховых дрожжей отличается тем, что большинство их совершенно не сбраживает раффинозу и лишь некоторые виды могут сбраживать раффинозу только на одну треть.
Низовые дрожжи во время брожения всегда садятся на дно и не образуют шапки на поверхности бродящего сусла. Но своей бродильной способности они отличаются от верховых дрожжей тем. что полностью сбраживают раффинозу.
В пивоварении наиболее широко применяются дрожжи низового брожения. Дрожжи верхового брожения употребляют значительно реже, главным образом для темных или специальных сортов пива.
Для специальных сортов пива, например английского портера, применяются также дрожжевые грибки, относящиеся к семейству торул (из рода слабобродящих бретаномицетов). Эти группы торул придают пиву особый своеобразный аромат. Они вводятся в пиво не в начале брожения, а в конце, перед выдержкой пива в лагерных подвалах.
Низовые дрожжи, применяемые в пивоварении, подразделяются на расы, отличающиеся по характеру роста на питательных средах (колонии), размеру и форме клеток, степени сбраживания сахаров, образованию ароматических веществ и т. д.
Все низовые дрожжи сбраживают глюкозу, фруктозу, сахарозу, мальтозу, раффинозу, не сбраживают инулин, лактозу и декстрины. Верховые дрожжи легко образуют споры, низовые же значительно труднее, а у некоторых рас пивных дрожжей вовсе не удается получить спорообразования.
Дрожжевые клетки низовых и верховых дрожжей имеют в большинстве случаев овальную форму. Форма клеток довольно изменчива; изменчив также в широких пределах и размер клетки. Поэтому в одной и той же чистой культуре дрожжей можно встретить то более круглые, то более вытянутые клетки.
Размеры клеток в разных культурах дрожжей колеблются от 6-8 до 12 мк по большему и меньшему диаметру клетки.
В клетках пивных дрожжей, как и всех других, различаются отчетливо выраженные оболочка, протоплазма с различными органическими включениями, больших или меньших размеров вакуоли и ядро.
Ядро в дрожжевой клетке обычно видно при специальных методах окрашивания. Оно имеет изменчивую форму, при почковании делится, и отделившаяся половина переходит в почку, из которой формуется новая, дочерняя дрожжевая клетка. При спорообразовании ядро клетки делится на столько частей, сколько спор образуется в клетке.
Морфологическое строение клетки зависит от ее физиологического состояния. Голодающие клетки, находящиеся в осадках, имеют зернистое строение протоплазмы и кажутся съежившимися. В мертвых клетках протоплазма отстает от оболочки, сами клетки кажутся сморщенными и имеют явно неправильное очертание оболочки. Такие клетки легко и интенсивно окрашиваются красками, например метиленовой синью. Молодые, только что отпочковавшиеся клетки имеют более нежную, тонкую протоплазму, в некоторых местах которой видны более темные, сильно преломляющие свет включения - жировые капельки, волютин и другие включения запасных питательных веществ. По мере старения клеток в них появляются больше включений и более выраженная зернистость протоплазмы, вакуоль также становится более заметной и большего размера.
Различное морфологическое строение клеток в зависимости от физиологического состояния дрожжей является одним из показателей, по которым можно при известном навыке судить ѳ благоприятных или неблагоприятных условиях брожения сусла.
Наиболее распространенной расой низовых дрожжей в пивоваренном производстве СССР была раса 776.
За последние годы широкое распространение получили также расы 11 и 47, выведенные Центральной научно-исследовательской лабораторией бродильной промышленности РСФСР.
Низовые пивные дрожжи обладают свойством во время брожения, и особенно в конце его, образовывать хлопья, состоящие из массы отдельных клеток, как бы склеенных между собой. Эта способность дрожжей (хлопьевидность) имеет весьма важное практическое значение. Благодаря хлопьевидности пивных дрожжей достигается быстрое осветление пива и создается полная возможность собирать дрожжи из бродильных чанов, не прибегая к сепарированию, и вновь использовать их для последующих циклов брожения.
Хлопьевидность дрожжей зависит от свойств оболочек дрожжей и также, по-видимому, от электрического заряда клеток. Она легко нарушается при изменении концентрации водородных ионов. В слабощелочной среде хлопьевидные дрожжи становятся пылевидными, но особенно сильно изменяется хлопьевидности дрожжей в кислой среде.
Наибольшая хлопьевидное™ дрожжей наблюдается в среде при pH 4,0-4,4. При большем подкислении среды дрожжи становятся пылевидными.
На характер хлопьеобразования дрожжей большое влияние оказывает температура брожения. При более высокой температуре хлопьеобразование менее выражено, чем при низких температурах.
На скорость оседания дрожжей известное влияние оказывает также упитанность дрожжей и их плотность. В этом отношении низовые дрожжи также выгодно отличаются от верховых, более легких дрожжей.
Оптимальная температура для роста низовых дрожжей около 25-27°С, минимальная - около 2-3°С. Низовые дрожжи хорошо переносят низкую температуру при хранении; при длительном хранении в этих условиях они приспосабливаются и значительно повышают бродильную энергию.
Дрожжи низового брожения при температуре 60-65ºC отмирают. Это имеет важное значение при пастеризации пива. Оптимальной температурой брожения в производстве обычно считается от 8 до 10ºC, хотя при более высокой температуре (10-14°С) может быть достигнута наибольшая интенсивность брожения.
На низовые дрожжи оказывает сильное угнетающее действие ряд веществ. При содержании этилового спирта в среде выше 1,5% замедляется размножение дрожжей, а при содержании спирта выше 3% заметно ослабевает их бродильная способность. Кислоты действуют угнетающе в значительно меньших концентрациях, причем минеральные кислоты действуют сильнее органических. Серная кислота в концентрации 0,5% убивает дрожжи в течение 1-2 ч. Из органических кислот молочная кислота в концентрации до I % легко переносится дрожжами. Сильное действие на дрожжи оказывает уксусная кислота. В концентрации 0,5% она резко угнетает, а в концентрации 1% убивает дрожжи в короткий срок.
Самая слабая органическая угольная кислота (Н2СО3) также угнетает развитие дрожжей. Это угнетающее влияние углекислоты при концентрации ее в среде до 0,2% сказывается главным образом на замедлении почкования дрожжей. Более высокая концентрация угнетает и бродильную способность дрожжей. Концентрация углекислоты 1,5%, достигаемая при избыточном давлении CO2 9-10 атм, останавливает брожение, но не убивает дрожжей.
Угнетающее действие угольной кислоты связано с торможением реакции расщепления пировиноградной кислоты на ацетальдегид и углекислоту (см. приведенную выше схему брожения).
Ацетальдегид, как и формалин, оказывает на дрожжевые клетки сильное угнетающее действие при относительно низких концентрациях, но более высоких, чем это встречается в нормальных условиях брожения.
Высшие спирты, в том числе и образуемые дрожжами при брожении, входящие в состав сивушного масла, ядовиты для дрожжевых клеток. Но в тех концентрациях, которые встречаются в бродящем пиве, заметного влияния на бродильную энергию не оказывают. Однако если искусственно создать такую концентрацию высшего спирта в среде до начала брожения, то может затормозиться или полностью остановиться процесс размножения и почкования дрожжей.
Ультрафиолетовые лучи обладают сильным действием и довольно быстро убивают дрожжи.
От свойства дрожжей зависят осветление пива во время брожения, степень сбраживания и скорость процесса брожения, а также аромат и вкус пива.
По способности обусловливать аромат и вкус пива дрожжи довольно резко отличаются друг от друга, хотя и не удалось еще химико-аналитическими методами установить различие, вызываемое ими в составе пива. Известно сейчас только одно, что эти различия зависят от продуктов обмена веществ у дрожжей, которые не удается определить химически, по всей вероятности, из-за крайне малых количеств этих продуктов.
Для производства пива наиболее ценны такие дрожжи, которые глубоко и быстро сбраживают экстракт сусла, дают сильное II полное осветление, образуют ярко выраженный аромат и мягкий вкус пива.
Однако такие качества дрожжей не совмещаются в какой- либо одной расе. Поэтому в производстве начинают широко применять смешанные расы дрожжей или ведут брожение на разных расах с последующим смешиванием пива после главного брожения. Для смешанных рас подбирают дрожжи с одинаковой скоростью размножения: одни с более выраженной бродильной способностью, а другие - дающие более тонкий аромат. Но при всех условиях смешиваемые расы дрожжей должны обладать примерно одинаково выраженной хлопьевидностью.
Все сказанное относится к культурным дрожжам. Кроме них, в производстве, как спутники и вредители, встречаются так называемые дикие дрожжи. Они могут относиться к тому же семейству зндомицетов и входить в группу сахаромицетов, как и культурные пивные дрожжи. Нередко на производстве встречаются дикие дрожжи, относящиеся и к другим семействам.
Из сахаромицетов встречаются дрожжи Saccharomyces еllі рsoideus, Saccharomyces pasteurianus, Saccharomyces apiculatus и др. Они обладают очень слабой энергией брожения, образуют не более 1% спирта. Многие из них бывают причиной постороннего запаха (затхлый, запах сельдерея) и вкуса пива.
К вредителям производства относятся также дикие пленчатые дрожжи (Mycoderma). Они преимущественно развиваются на поверхности сусла и пива в виде пленки, образуют вещества, придающие пиву неприятный вкус и запах, спирт окисляют до уксусной и угольной кислоты.
Большинство диких дрожжей нуждается в кислороде и довольно плохо развивается при температуре ниже 10°С. Поэтому предупредительные меры, предохраняющие производство от развития диких дрожжей, сводятся к соблюдению чистоты, ведению брожения при низкой температуре, уменьшению поверхности соприкосновения сусла и бродящего пива с воздухом.

 Калорийность Морская капуста (ламинария)
Калорийность Морская капуста (ламинария) Виноградное вино в домашних условиях: технология изготовления
Виноградное вино в домашних условиях: технология изготовления Как вкусно пожарить щуку на сковороде – полезные секреты приготовления и лучшие рецепты Щука рецепты приготовления на сковороде
Как вкусно пожарить щуку на сковороде – полезные секреты приготовления и лучшие рецепты Щука рецепты приготовления на сковороде Куриные ножки с рисом в духовке
Куриные ножки с рисом в духовке