제빵 효모의 인종 및 계통. 와인 효모의 인종 간의 차이점. 맥주 경주 및 균주의 일반적인 특성
섹션 3 맥주 생산.
6장. 맥주 맥아즙의 발효 .
양 조용 효모.
효모 세포 구조... 효모는 유대류 균류에 속하는 단세포 유기체입니다. 효모 세포의 모양은 타원형이며 원형이며 타원형입니다.
효모 세포에는 세포막이 위치한 세포벽 1이 있습니다. 막은 세포와 환경 사이의 대사에 영향을 미치는 선택적 투과성을 가지고 있습니다.
예를 들어, 아미노산과 포도당 분자는 더 작은 금속 이온보다 빠르게 막을 통과합니다. 세포는 이중막으로 둘러싸인 원형 또는 타원형 핵 2를 포함합니다. 핵소체는 핵 내부에 있습니다. 핵은 성장과 성장을 보장하는 대사 과정에 필요합니다.
효모 번식.
세포의 기초는 세포질 3이며 점성이 있고 약간 황색을 띤 액체입니다. 그것은 많은 기능을 수행합니다. 예를 들어 호흡의 첫 번째 단계와 알코올 발효는 세포질에서 직접 발생합니다. 다음은 세포의 구조적 요소입니다: 액포 4, 미토콘드리아 5, 리보솜 6. 미토콘드리아는 산화 대사와 관련된 과정이 일어나는 매우 작은 물방울 모양의 입자입니다. 리보솜은 막으로 둘러싸인 소포입니다. 리보솜은 단백질 합성이 일어나는 곳입니다. 액포는 세포액으로 채워져 있고 액포막에 의해 세포질과 분리된 공동입니다. 그들은 효모 세포의 성장과 번식을 일으키는 메타 크로마틴을 함유하고 있습니다. 산화 환원 과정은 액포에서 발생합니다.
효모 세포의 크기는 인종, 효모의 생리학적 상태 및 영양 배지의 조성에 따라 다릅니다. 압축 효모는 약 30%의 건조 물질과 70%의 수분을 함유하고 있습니다. 효모의 건조물은 90-95%의 유기물과 5-10%의 무기물을 포함합니다. 유기 물질 중에는 단백질 및 질소 함유 물질 - 54-56%, 탄수화물 - 24-40%, 지방 - 2-4%(건조 물질의 중량 기준)가 있습니다. 탄수화물의 주요 부분은 전분 아밀로펙틴과 화학 구조가 유사한 글리코겐(저장 물질)으로 표시됩니다. 무기물 중 인산의 약 절반, 칼륨의 약 1/3.
효모 재는 다음을 포함합니다(%): Р 2 О 5 -47-53, К 2 О-28-40; CaO-0.4 -11.3; Mg O-3.0-7.4, SiO2-0.28-0.73, SiO3-0.09-0.74; Сl -0.1-0.65. 게다가 에 소량 S, Zn, Mn, Cu, Fe가 있습니다.
인 화합물은 알코올 발효의 중간 물질의 일부이기 때문에 효모 세포의 대사에 중요하고 칼륨은 단백질 및 탄수화물 분자 구성에 주요 역할을 합니다. 효모는 비타민 B가 풍부하고 에르고스테롤(프로비타민 D) 등을 함유하고 있습니다. 효모에는 가수분해 및 합성 과정, 발효 및 호흡 과정과 관련된 다양한 효소 시스템이 포함되어 있습니다.
효모 성장 단계... 효모 성장을 효모 세포 수의 증가라고 합니다. - 재생산. 정상적인 조건에서 효모 세포는 출아에 의해 증식합니다. 모세포는 신장을 형성하여 딸세포로 성장합니다. 영양소가 부족하거나 기타 불리한 조건이 있으면 세포 내부에 칸막이가 형성되고 이러한 칸막이를 따라 세포가 분해되어 포자를 형성합니다. 가 있는 환경에서 좋은 조건영양 포자는 발아하여 새로운 효모 세포를 형성합니다. 맥주 맥아즙에는 세포 증식에 필요한 모든 물질이 포함되어 있으므로 맥아즙을 발효할 때 효모는 포자를 형성하지 않고 발아에 의해서만 번식합니다.
효모를 맥아즙에 도입한 후 양적 및 질적 변화가 관찰됩니다. 효모의 양은 여러 번 증가하지만 분산 상태의 농도가 먼저 증가하여 최대 값에 도달한 다음 감소합니다. 맥주 맥아즙의 발효 중 효모의 번식은 여러 단계로 진행됩니다. 성장 곡선에서 여러 단계를 구분할 수 있습니다. (세로 - 효모 세포의 수, 가로 - 시간.)
 잠복기 또는 지연기(성장 지연)라고 하는 초기 단계에서 효모는 새로운 환경에 적응하고 번식을 위해 스스로 준비합니다. 이 단계는 일반적으로 세포가 환경에 적응하는 실제 휴식 단계와 번식이 점진적으로 시작되는 단계의 두 부분으로 나뉩니다. 맥주 효모의 잠복 기간은 1-1.5일입니다. 그 안에 세포의 부피가 증가하고 길어지며 신진 세포의 비율이 증가합니다.
잠복기 또는 지연기(성장 지연)라고 하는 초기 단계에서 효모는 새로운 환경에 적응하고 번식을 위해 스스로 준비합니다. 이 단계는 일반적으로 세포가 환경에 적응하는 실제 휴식 단계와 번식이 점진적으로 시작되는 단계의 두 부분으로 나뉩니다. 맥주 효모의 잠복 기간은 1-1.5일입니다. 그 안에 세포의 부피가 증가하고 길어지며 신진 세포의 비율이 증가합니다.
대수라고 하는 다음 단계에서는 효모 증식 속도가 최대이고 모든 세포가 활성화되고 발효 배지에 부유됩니다.
대수 단계 후, 세포 증식이 느려지고 사망 및 번식 속도가 균형을 이루는 동안 정지기가 시작되어 결과적으로 살아있는 세포의 수가 변하지 않습니다.
부패 단계라고 하는 마지막 단계는 영양소의 질량 감소와 대사 산물의 양 증가로 인한 세포 활동의 감소를 특징으로 합니다. 번식이 멈추고 세포가 죽어 발효 장치의 바닥에 가라앉습니다.
살아있는 효모 세포에서 중요한 활동은 다양한 생화학적 과정에 의해 지원되며, 죽으면 이러한 과정의 일관성이 파괴되고 자가 분해가 시작됩니다. 자체 효소의 영향으로 세포가 부패합니다. 이 경우 세포의 구조가 교란되고 일부 효소의 활성이 증가하고 다른 효소의 활성이 약화됩니다.
예를 들어, 가수분해 효소는 활성화되고 호흡 및 발효 효소는 죽습니다. 효모의자가 분해 중에 단백질 물질, 탄화수소, 지방, 유기 인 화합물이 분해되고 저분자 분해 생성물이 형성되어 세포벽을 통해 맥주로 확산되어 맛이 바뀝니다. 약간의 자가 분해에서는 약한 효모 향이 나타나고 강한 자가 분해에서는 쓴맛이 나는 이질적인 맛이 나타납니다. 자가 분해 중에 방출되는 질소 물질은 맥주의 콜로이드 헤이즈일 수 있습니다.
맥주 효모의 인종.양조 산업에서는 Saccharomycetaceae 계통과 Saccharomyces 속에 속하는 문화 효모만 사용됩니다. 하면발효효모 Saccharomyces carlsbergensis와 상면발효효모 Saccharomyces cerevisiae가 있습니다.
처음에는 발효가 상온에서만 일어나기 때문에(와인 양조, 베이킹에서와 같이) 상면 발효 효모가 알려졌습니다. 이산화탄소로 포화된 음료를 얻기 위해 그들은 낮은 온도에서 발효되기 시작했습니다. 변경된 외부 조건의 영향으로 특정 특성을 가진 하면 발효 효모가 얻어졌습니다.
양조에서는 하나 이상의 기능이 서로 다른 효모 품종이 사용됩니다. 그들은 하나의 세포에서 얻습니다. 그러한 문화를 종족(계통)이라고 합니다.
집중발효과정에서 상면발효효모는 발효액의 표면에 떠서 거품층의 형태로 축적되어 발효가 끝날 때까지 이 형태를 유지한다. 그런 다음 발효 장치의 바닥에 매우 느슨한 층을 형성하여 침전됩니다. 이러한 효모는 그 구조에 의해 가루 효모로 분류되며, 응집성 풀뿌리 효모와 달리 서로 달라붙지 않습니다.
하면발효효모는 맥주거품의 표층으로 들어가지 않고 발효장치의 바닥에 빠르게 가라앉는다.
효모가 응집하는 능력은 맥주의 정화를 가속화하고 발효 후 발효 장치에서 효모를 제거하고 종자 효모로 후속 사용을 촉진하기 때문에 맥주 맥아즙 발효 기술에 중요합니다. 낮은 발효 온도는 응집을 촉진합니다.
배지의 반응은 효모의 특성에 큰 영향을 미칩니다. 예를 들어, pH 3 미만의 산성 환경 및 pH 8 이상의 알칼리성 환경에서 응집 효모는 분쇄된다. 플레이크 효모는 분쇄 효모에 비해 세포가 더 크고 자가 분해에 덜 민감하며 바이오매스가 크게 증가하고 발효 활성이 낮고 맥주에서 디아세틸과 고급 알코올을 덜 형성하여 품질에 긍정적인 영향을 미칩니다. .
하면발효효모는 라피노스를 완전히 발효시킨다는 점에서 상면발효효모와 다릅니다. 하면발효 효모는 최적의 성장 온도가 25-27C, 최소 2-3C이며 60-65C에서 죽습니다. 다운스트림 효모의 최대 발달은 pH 4.8-5.3에서 발생합니다. 맥아즙에 용해된 산소는 효모의 성장을 촉진하는 반면 발효 생성물(에틸 알코올, 이산화탄소, 고급 알코올, 아세트알데히드, 산)과 증가된 당 농도는 효모의 성장을 억제합니다.
맥주 효모는 다음 요구 사항을 충족해야 합니다. 맥아즙의 빠른 발효, 발효 중 맥주의 우수한 플레이킹 및 정화로 맥주에 깨끗한 맛과 쾌적한 향을 제공합니다.
효모의 발효 활성은 맥아즙의 발효 정도에 따라 결정됩니다. 발효도(V)는 초기 맥아즙(E)의 건조 물질 질량에 대한 발효 추출물(Ee)의 질량 비율을 나타내는 백분율로 표시되는 지표입니다. V = ((Ee) 100 ) / E, 여기서 e는 맥주의 추출물 함량, 맥주의 중량%입니다.
발효 정도에 따라 강발효 또는 고발효(발효도 90~100%), 중발효(80~90%), 약발효, 저발효(80% 미만)로 구분한다.
효모 인종은 강력하게 발효됩니다: 11, F(체코 공화국에서 구입), 균주 8a(M). 레이스 11 효모는 원료의 품질에 구애받지 않고 잘 정착하며 맥주의 풍미가 가득한 것이 특징입니다. 레이스 F의 효모는 맥주를 밝게 하고 기분 좋은 향을 내며 감염에 강합니다. 효모균주 8a(M)는 발효활성이 높고 배율이 높아 잘 가라앉는데 이 효모를 사용하면 본발효기간을 7일에서 5일로 단축하여 맛이 좋은 맥주를 얻을 수 있다.
중간 발효 효모에는 776, 41, 44, S(Lviv), P(체코 공화국),
A(리가). 효모 레이스 776은 원료에 소박하며, 맥아가없는 재료를 사용하여 맥주를 양조하는 데 사용할 수 있습니다. 완성된 맥주는 맛이 만족스럽고 홉의 쓴맛이 강하다. 효모 레이스 41, 44, S, 침강력이 좋고 맥주의 맛이 깨끗하고 순하며, 효모 레이스 44는 경도를 높인 물을 사용하면 좋은 맥주를 얻을 수 있습니다. F, A 인종의 효모는 맥주를 잘 정화하고 감염에 강합니다.
상면 발효 효모는 흑맥주 및 특수 맥주에 사용됩니다.
효모의 품질에 대한 요구 사항이 항상 한 인종에 의해 충족되는 것은 아니므로 생산에 여러 인종의 혼합물을 사용하거나 다른 인종에서 맥아즙을 별도로 발효한 다음 젊은 맥주를 혼합합니다.
순수 배양 효모 사육.
육종은 한 시험관의 질량에서 발효 장치에 도입하는 데 필요한 모액의 질량으로 효모의 질량이 증가하는 것을 의미하는 것으로 이해됩니다.
전체 번식 과정은 실험실(실험실에서 효모 번식)과 작업장(순수 배양 부서에서 효모 번식)의 두 단계로 구성됩니다.
실험실 단계는 여러 개의 연속적인 구절로 구성됩니다. 먼저, 시험관의 순수한 배양물을 멸균 맥아즙에 원뿔 모양으로 계대한 다음, 멸균 발효 맥아즙이 있는 효모를 새로운 멸균 맥아즙에 계대 배양합니다. 그 부피는 계대 배양에서 계대 배양으로 몇 배 증가합니다. 실험실 단계는 7-8 ° C에서 5-6 일 동안 Carlsberg 구리 플라스크에서 6 리터의 맥아즙 발효로 끝납니다.
작업장 단계는 특수 장치에서 멸균 된 호프 맥아즙에 효모를 번식시키는 것입니다.
그림은 작업장에서 순수 효모 배양을 위한 Greiner 식물을 보여줍니다(파이프라인은 표시되지 않음). 설비는 멸균기(4), 사용된 효모의 양에 따라 개수가 달라지는 2개의 발효 실린더(3), 사전 발효 탱크(1) 및 종자 효모용 용기(2)로 구성됩니다.
살균기 및 사전 발효 탱크에는 맥아즙 가열 또는 냉각 코일, 공기 필터 및 기기가 장착되어 있습니다.
발효 실린더에는 10리터 용량의 종자 효모용 용기가 있습니다.
살균기는 맥아즙 끓이기 (살균) 및 후속 냉각, 발효 실린더-전파의 첫 번째 단계, 사전 발효 탱크-맥즙의 살균 및 냉각 및 전파의 두 번째 단계를 위한 것입니다. 순수한 문화의. 대기 온도 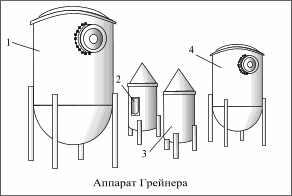 순수 문화부 8-9 о С.
순수 문화부 8-9 о С.
순수한 문화를 육성하는 것은 다음과 같습니다. 멸균기 4에서 뜨거운 호핑된 맥아즙을 맥즙 보일러에서 수집하고 1시간 동안 끓인 다음 8-12°C로 냉각합니다. 압축된 멸균 공기를 사용하여 맥아즙을 실린더 3에 공급합니다. 칼스버그 구리 플라스크의 밸브를 닫은 다음 3일 이내에 발효하는 동안 효모가 증식하고 질량이 증가합니다. 3일이 지나면 사전 발효 탱크에 맥아즙을 채우고 끓일 때까지 가열한 다음 냉각합니다. 발효 실린더 3의 순수한 배양액의 일부는 종자 효모용 용기 2에 저장하기 위해 가져와 다음 희석까지 저장하고, 주요 부분은 9°C에서 예비 발효가 수행되는 저장소 1로 펌핑합니다. 3일 동안.
다음 희석 주기에서 발효 실린더 3의 멸균 맥아즙에 접종하기 위한 효모는 용기 2에서 가져옵니다. Greiner 설비에서 순수 배양물을 희석하는 과정은 효모에서 외부 미생물총이 감지될 때까지 여러 번 반복됩니다.
저장소 1의 발효 가능한 덩어리는 1000 dal 용량의 예비 발효를 위한 특수 장치로 펌핑되지만 5-7 o C의 온도에서 1/3 맥아즙으로 채워집니다. 발효 12시간 후, 또 다른 400 dal의 신선한 호핑된 맥아즙을 이 장치에 첨가하고 온도를 5-7C로 유지하면서 추가로 36시간 동안 발효를 계속합니다. 그런 다음 발효된 맥아즙은 700달의 맥아즙에서 주요 발효 장치로 펌핑되고 1일 후에 맥즙으로 가득 차고 온도, 맥아즙 농도 및 정화를 제어하면서 일반적인 방식으로 발효가 수행됩니다. 발효 과정에서 침전된 효모는 씻어내고 찬물로 씻어 1세대로 생산에 사용한다.
작업을 시작하기 전에 효모 재배 장치를 0.15-0.17 MPa의 압력에서 45 분 동안 증기로 멸균합니다. 살균기로 들어가는 공기는 공기 필터를 통과해야 합니다.
알코올 발효- 와인, 위스키, 맥주 등 모든 알코올 음료의 기초이자 시작입니다. 바로 이 기초의 기초는 원료, 물 및 효모입니다. 이 기사에서는 다양한 유형에 대해 이야기 할 것입니다. 와인 효모국내 및 산업용 포도주 양조에 사용됩니다. 어떤 종류의 효모가 있는지 생각해 봅시다. 친숙하고 와인의 풍부함과 다양성을 개발하는 데 도움이되며 와인 메이커에 적대적이며 와인 자체를 억압하고 손상시킬뿐만 아니라 장비와 함께 전체 와이너리를 감염시킵니다.
알코올 발효("발효"라고도 함)는 효모에 의해 수행되는 생화학적 과정으로, 이상적인 결과는 당류(주로 자당, 포도당 및 과당)를 에틸 알코올(주요 제품), 이산화탄소 및 많은 것으로 전환하는 것입니다. 화학적 미량 원소(필수 및 필요하지 않음, 유해하고 유익한 부산물).
누룩- 미세한 단세포 곰팡이. 현대 미생물학은 그것들을 150만 종과 수천 개의 아종으로 나누며, 이미 그들은 통제되거나 통제되지 않은 돌연변이와 재생의 결과에 따라 많은 변이를 재현할 수 있습니다. 하나의 동일한 효모를 여러 번 사용하여 독립적으로 전파합니다).
고대부터 사람은 알코올 발효를 채택했지만 식품 산업과 과학은 여전히 에틸 알코올 생산에 효모를 사용하는 새로운 가능성과 특징을 점점 더 많이 열어주고 있습니다. 많은 노력이 포도주 양조 시장 및 이와 관련된 미생물학의 발전에 정확하게 집중되어 있습니다. 이것은 전체 산업 - 양조학입니다. Oenology는 박테리아 연구 및 번식, 효소 개발, 와인 메이커에 필요한 특성을 지닌 효모 연구 및 번식, 많은 와인 및 와인 음료 생산 허용, 새로운 측면과 맛 발견, 보존뿐만 아니라 인류의 역사적 유산이 된 오래되고 희귀한 것들.
생산에 사용되는 주요 유형의 효모(Saccharomycetes - 그들은 모든 알코올 음료(와인 메이커, 양조업자 및 밀주업자)의 친구입니다) 알코올 음료(집에서 포함).
명확성을 위한 표입니다. 다음은 산업(일부 국내) 포도주 양조법에서 인기 있는 몇 가지 효모 인종입니다. 특정 종족의 선택은 무엇보다도 발효 조건에 기인한다는 점에 유의하십시오. 포도주 양조에 권장되는 포도주 효모의 일부 인종이 표에 나와 있습니다(일부 외국 제품은 저희 가게에서 구입할 수 있습니다).
"사카로마이세스 세레비지애"(Saccharomyces cerevisia)는 현재 세계에서 가장 광범위하고 다양하며 "길들인" 효모입니다. 베이커리, 와인, 맥주 및 알코올 효모... 그것들은 매우 다양하고 적용 영역이 넓기 때문에 별도의 기사가 필요합니다. 불행히도, 그들은 야생 포도주 양조법에서 항상 발견되는 것은 아니며 Cerevisia의 Saccharomycetes 중 와인에 부정적인 영향을 미치는 수백 개의 아종이 있다는 점을 감안할 때 "성공적인 감염"의 확률은 훨씬 낮지 만 존재하지 않습니다. ;
"사카로마이세스 비니"(Sacharomyces vin) - 그들은 주로 잘 익은(특히 손상된) 포도와 주스(외부 영향을 받지 않음)에서 삽니다. 그들은 종종 토양, 곤충의 소화 시스템(특히 초파리, 말벌 및 꿀벌)과 와이너리 내부(가정 와이너리 포함) - 벽, 접시 및 장비에서 찾을 수 있습니다. 그러나 가정용 포도주 양조법에서는 매우 제한적으로 사용되며 많은 바람직하지 않은 영향을 줄 수 있습니다. 예를 들어 포도주의 흐림 및 현탁액 형성;
"사카로마이세스 오비포르미스"(Saccharomyces oviformis) - 대부분의 경우 이 품종을 포도주 양조에 사용하면 유익할 수 있습니다. 그들은 당도가 높은 맥아즙을 발효시키는 데 사용되며 드라이 와인 생산에 매우 적합합니다. 이 효모 인종의 현대 대표는 샴페인 생산에 인기가 있습니다.
"Leningrad", "Kiev"와 같은 국내 인종도 있습니다. 이 균주를 사용할 때의 단점은 탁도 및 늦은 침전물의 형성뿐만 아니라 완성된 와인의 발효 재개(가장 흔히 반단 맛이지만 뿐만 아니라)를 포함할 수 있습니다. 셰리 주정 강화 와인 생산을 위해 이러한 균주를 가장 생산적으로 사용합니다. 이 때문에 투옥된 대표자(" Oviformis Cheresiensis") -" Jerez 96-K "및" Jerez-20-S "- 그러나 강한 와인(16-17% vol.)에 대한 필름을 매우 빠르게 생성합니다.
"사카로마이세스 바야누스(우바룸)"(Saharomyces bayanus uvarum) - 대부분 과일 와인과 주스에서 찾을 수 있습니다. 이것은 매우 서두르지 않은 효모입니다. 천천히 발달하고 현재의 통제 수준으로 구별하기 어려운 가정용 포도주 양조의 미생물 학적 과정으로 인해 돌연변이를 통제하기 어렵습니다. 그들은 높은 수준의 감쇠(알코올 생산)를 가진 효모가 아니지만 안정성과 추위에 대한 저항이 증가하는 드문 특징이 있습니다. 발효산물은 앞서 언급한 효모균주와 거의 동일하다. 에스비니... 특징 -이 품종의 많은 (전부는 아님) 인종은 가장 밀도가 높은 (교반에 취약하지 않은) 효모 침전물을 형성 할 수 있으며 거품이 거의 완전히 없으며 글리세린 함량이 증가합니다. 가장 인기있는 종족 - "Novotsimlyanskaya 3"은 거의 액세스 할 수 없지만 가정에서 포도주 양조법, 그러나 세미 스위트 와인 생산에 탁월한 것으로 입증되었습니다.
그러나 모든 효모가 똑같이 만들어지는 것은 아닙니다. 다음으로 와인 메이커의 위험하고 비열한 몇 가지 적, 즉 매우 비우호적 인 특성을 가진 효모를 고려하십시오.

"피키아" / "한세눌라" / "칸디다" 및 기타 더러운 것들 - 심각한 적, 해충 및 실패한 와인의 범인(특히 사용 시 야생 효모). 그들의 주요 특징은 특히 호기성 조건에서 와인 표면에 필름이 형성된다는 것입니다 ( 냄새 트랩(워터 씰) 도움). 이 해로운 효모의 세포는 불안정한 모양을 가지고 있습니다. 타원형, 타원형, 소시지 및 클럽 모양이 있으며 불균형하게 길쭉합니다. 그들 중 일부(Pichia 및 Hansenula)는 포자를 형성하는 반면 다른 일부는 출아로 번식합니다. 이 종족은 맥아즙을 빠른 속도로 발효시켜 산화시킬 수 있습니다. 그들 중 일부는 Hansenula와 같은 현대 와인에 충분한 알코올을 축적할 수 없습니다. 최대 5%의 에탄올만 제공합니다.
적절하게 준비된 맥아즙에서는 일반적으로 위험하지 않습니다. 소량 함유. 와인이 만들어지는 와인 재료의 준비 및 저장을 위한 기술 조건(밀접함, 재료의 무균성 및 주변 분위기)을 따른다면 두려울 것이 없습니다. 그러나 준비가 좋지 않으면 (초보자 / 포도를 씻지 않는 것을 좋아하는 무지한 와인 메이커 (미생물을 알지 못함), 처리되지 않은 물과 용기 사용) - 변형이 표면에 빠르게 번식하기 시작합니다. 이로 인해 빠르면 2-3일(때로는 나중에)에 광택 필름이 생성되고 접힌(여드름이 있는) 필름이 생성됩니다. 이 필름은 흰색과 흐릿한 투명에서 회색 색상에 이르기까지 다양한 색상 범위를 가질 수 있습니다.
와인이 갑자기 오염되면 필름이 최악이 아닙니다.필름 형성은 발효의 새로운 단계인 설탕의 산화를 통한 발효를 나타냅니다. 이 과정에서 에틸뿐만 아니라 아밀 및 부틸 알코올뿐만 아니라 아세트산, 부티르산, 숙신산 및 이러한 산의 다양한 에스테르 화합물이 반드시 형성됩니다. 결과적으로, 맥아즙은 특징적인 매우 불쾌한 냄새를 얻습니다. 펄프를 사용하는 경우 특히 가능성이 높습니다. Pichia / Hansenula와 같은 인종에 가장 유리한 조건이 옵니다. 프로세스).
일부 필름형 효모(종종 야생 발효에서 발견됨 - 반대로 매우 내알코올성(또한 아황산염 내성)가 있을 수 있으며, 처음에는 일부 불운한 포도주 양조업자를 기쁘게 합니다. 그들은 알코올 함량이 14% vol. 그리고 400-500 mg / l SO2 같은 이유로 테이블 와인 (산소를 사용할 수 있음)에서는 매우 편안하게 느껴지고 점차적으로 와인을 부패시키는 다양한 불순물을 형성합니다 (특히 방부제가없는 수제 및 미완성 와인). 황산염, 아황산염, 황화수소 및 각종 황화합물을 많이 발생시키면서 불쾌한 썩는 냄새의 원인이 됩니다.
Zwel은 종종 맥아즙에 앞서 언급한 인종 및 품종의 활성 효모의 존재로 인해 불쾌한 외관(강수, 부유, 섬, 탁도)과 불쾌한 냄새를 주는 가장 흔한 와인 질병 중 하나입니다. 또한, 그들은 드라이 와인, 샴페인 및 셰리의 최악의 적입니다. 조심하십시오!
와인에서 악성 효모의 발생 및 번식은 산소 접근을 제한하고(다시 말하지만, 물개) 와인 재료를 저장하기 위한 올바른 조건(저온 및 용기 자체를 완전히 채워야 함)을 설정하여 방지할 수 있습니다. 많은 여유 공간을 남기지 않고. 때로는 제때에 충전해야 할 수도 있습니다. 이를 무시하지 마십시오.
"접합당류"- 해로운 (와인 메이커를 위한) 미생물총의 또 다른 대표자. 이 효모 종족은 삼투압성이 매우 높아 극한의 날씨에서도 번성합니다.차 달콤한 wort는 100ml 당 60 및 80g의 설탕 함량으로도 개발됩니다. (wort의 진공 발효, 다양한 수제 제품 - 꿀, 잼, 보존 식품). 그들은 그러한 극한 조건에서 발효 과정을 일으키고(이것이 얼마나 놀라운 순환수 모듈인지 상상해보십시오) 제품을 근본적으로 손상시킵니다. 역설적으로, 이 균주는 내알코올성이 매우 낮습니다. 평균적으로 10-11% 이하의 알코올을 형성하고 매우 오랜 시간 동안 발효되지만 이 시간 동안 한때 적합했던 재료/제품을 완전히 망칠 수 있습니다. 그러나 현대 과학은 "Races Vierul, May-kopskaya, Krasnodar 40"과 같이 제한적이긴 하지만 매우 신맛이 나는 맥아즙의 산도를 줄이는 데 사용할 수 있다는 점을 발견했습니다. (나머지 유해한 물질과 마찬가지로) 산화가 아닌 주로 발효에 의해 물질을 재활용합니다.
"사카로미코드"- 고대와 현대 포도주 양조의 다음 해충. 이것은 큰 모양과 불안정한 유형의 번식을 가진 복잡한 효모입니다. 분열하고 새싹이 나올 수 있습니다. 그들은 최대 12%(일부 보고에 따르면 최대 14%)까지 알코올에 저항하며, 맥아즙 자체에는 순수한 효모 배양물의 생존에 해로운 영향을 미치고 다양한 부작용을 일으킬 수 있는 에틸 아세테이트가 풍부합니다. 예를 들어, 발효의 갑작스런 재개).
"Saccharomycodes ludwigii"- 고도로 황화 된 맥아즙의 경우에도 발효 재개를 유발할 수 있습니다(모든 방부제가 만병 통치약은 아님). 또한 완성된 와인과 맥아즙의 매우 높은 SO2 함량을 견딥니다.
"한세니아스포라"(apiculatus) - sporogenes (Hanseniaaspora apiculata)와 asporogenic 균류 (Klocker apiculata)로 나타낼 수 있는 매우 흔한 효모. 과일이 손상되면 이미 거기에있을 가능성이 높으며 주스와 큰 과일을 경멸하지 마십시오 (작은 과일도 약간 덜 자주). 흥미로운 사실: 포도가 익으면 거기에 있는 모든 효모의 99%까지 도달할 수 있습니다(순도를 선호하는 또 다른 이유 - 포도와 모든 와인 재료를 씻으십시오!). 발효 능력은 낮습니다 - 약 4-7% vol. 그러나 알코올, 휘발성 화합물, 에틸 아세테이트, 부티르산, 프로피온산 및 기타 산이 매우 풍부합니다. 종종 그것들은 맥아즙이나 포도주(특히 집에서 만든 포도주)가 발효되지 않은 이유입니다. 그들은 폭발적으로 성장하고 맥아즙에서 매우 빠르게 번식하여 다른 효모, 특히 고귀한 효모의 성장을 여러 번 능가합니다. 그들은 와인의 쓴맛, 많은 외국과 동시에 강한 냄새와 식초 색조를줍니다. 수제 샴페인이나 셰리를 만들고 "끈적끈적한" 잔여물을 얻었습니까? 이것도 그들의 수작업입니다. 황산화(때때로 증가)와 장기간 침강이 도움이 될 수 있습니다.
"토룰롭시스"- 특히 포도와 관련하여 포도주 양조에 해로운 광범위한 인종. 그들은 야생 효모 (소위 고귀한 부패)로 시작된 이미 발효 된 포도 주스에서 적극적으로 배설됩니다. 그들은 두 가지 주요 유형(T. bacillaris 및 T. Candida)으로 나눌 수 있습니다. 최근 몇 년 동안 포자를 형성한다는 정보가 있었지만 현재는 출아로 번식하는 것이 일반적으로 받아들여지고 있습니다. 와인에는 자주 등장하지 않지만 포도 주스에는 자주 등장한다. 에틸 알코올 회전율의 최대 12.5%까지 맥아즙을 발효시킬 수 있습니다. 생화학 측면에서 야생 발효 중에 나타나는 다른 효모만큼 해롭고 파괴적이지 않으며 훨씬 덜 해로운 화학 원소를 형성하지만 점액을 형성합니다. 동의하십시오. 와인을 마시는 것을 기뻐할 사람은 거의 없을 것입니다. 일부 지역에서는 젤리 또는 더 끈적 끈적한 것과 유사한 섬을 발견합니다. 그들은 또한 삼투압체(당 함량이 100ml당 60-80g인 경우에도 기분이 좋음)이며 고온을 좋아하며 SO2의 증가는 전혀 눈에 띄지 않습니다.
"로도토룰라"- 이것은 소위 "핑크 효모"이며, 특징적인 색 때문에 그렇게 불립니다. 그들은 설탕을 발효시키지 않고 산화시켜 짙은 분홍색 막을 만듭니다. 주스의 산화, 탁도 형성 및 디저트 및 세미 스위트 (잘, 달콤한) 와인의 침전에 기여합니다. 흥미로운 특징은 공기 중의 알코올 증기를 먹을 수 있다는 것입니다. 이러한 이유로 그들은 종종 와이너리의 벽과 심지어 지하실의 분홍색 점액 형태로 "빨간 손에 잡혔습니다".
출력 대신:
생산 조건에서 자연 발효는 바람직하지 않은 결과를 초래할 수 있습니다. 이것을 피하고 좋은 품질의 와인을 얻기 위해 특별히 선택된 효모 균주의 순수한 배양물에서 발효가 수행되어 지시된 과정을 위해 맥아즙에 도입됩니다. 현대의 포도주 양조법에서는 여전히 중립적이고 포도주 양조에 적합한 효모를 발견하는 것이 가능합니다. 그것들은 사실이 아닐 수도 있지만(순수 효모 배양과 같이 이상적임), 포도주를 망치지는 않을 것입니다. 이러한 이유로 자연발효가 반드시 와인의 부패로 이어진다고 단정할 수는 없지만 이러한 가능성은 항상 존재하며 때로는 이 확률이 범주적으로 증가할 수 있습니다. 예를 들어, 포도 한 송이에서 유해한 킬러 효모가 발견되면 발효 중에 민감한 효모를 엄청난 속도로 파괴할 수 있습니다(1개의 킬러 세포는 평균 20개의 비활성 세포를 파괴할 수 있음). 또한, 일반적으로 악취가 나는 불필요한 화학 물질로 와인을 포화시킬 수 있는 악의적으로 중립적인(종내 투쟁에 참여하지 않는) 인종이 있습니다.
야생 효모의 사용은 종종 불친절함으로 가득 차 있습니다. 갑자기 맥아즙이 "끓는 것"을 멈추거나 "또 다른 발효"(필름 같은)가 시작됩니다. 따라서 야생 효모를 범주적으로 사용하는 경우 다음을 추가하십시오. 이산화황 , 고귀한 균주는 일반적으로 황산염에 내성이 있는 반면 유해한 효모는 일반적으로 그렇지 않습니다(그러나 불행히도 전부는 아닙니다)
모두에게 맛있는 와인!
당사에서 언급된 제품.
베이커리 효모 - 2
문학:
세미카토바 N.M. 베이커리 효모: - 모스크바: 식품 산업 출판사, 1980 - 200 p. 코드 6P8.2 C30, inv. 번호 845314 хр
Matveeva I.V., Belyavskaya I.G. 제빵의 생명공학적 기초. - M .: DeLi 인쇄, 2001-150 p. (유.)
온도의 영향.
온도에서 효모의 특정 성장률:
20 ° C = 0.149; 30 ° C = 0.311; 36 ° C = 0.342; 40 ° C = 0.200; 43 ° C = 0
영향환경의 활성 산도.
높은 성장률 제빵사의 효모 pH = 4.5 - 5.5에서 관찰됨. 제빵용 효모를 pH 3.0-3.5로 배양하는 동안 배지를 산성화하고 8.0으로 알칼리화하면 효모 세포의 증식이 중지되고 효모의 품질이 저하됩니다.
화학 물질의 영향.
배지의 함량이 (%) 이상인 경우 효모 성장이 억제됩니다. 아황산 무수물 - 0.0025, 불화 나트륨 - 0.002, 아질산염 - 0.0005, 포르말린 - 0.001, 카라멜 - 0.1.
효모의 성장은 또한 배지의 함량이 더 많을 때(%) 산에 의해 지연됩니다: 옥살산 - 0.001, 포름산 - 0.0085, 아세트산 - 0.02, 부티르산 - 0.005.
또한, 배지 내 함량이 0.02-0.1 이상(%)인 경우 상기 산의 염에 의해 효모의 성장이 억제된다. 약 0.1%의 산염 농도는 효모의 성장을 억제합니다.
금속염은 비소 - 0.0005, 구리 - 0.005, 은 - 0.000001과 같이 환경 내 함량이 (%) 이상일 때 파괴적으로 작용합니다. 금속염의 살균 효과는 온도 조건, 효모의 총 농도, 배지의 조성 및 산도에 따라 다릅니다.
효모 속도는 또한 0.004% 이상의 농도에서 질산염을 효모에 유독한 아질산염으로 환원시키는 박테리아에 의해 생성된 아질산염에 의해 억제됩니다.
항생제는 효모 활동을 감소시키지 않습니다.
효모 배양 중 환경 물질 농도의 영향.
효모 배양을 위한 영양 배지의 최적 당도는 5~6%입니다.
효모 세포의 삼투 감도는 매우 중요합니다. 즉, 증가된 농도의 염화나트륨(밀가루의 약 2중량%)에서 당을 발효시키는 능력입니다.
설탕이 포함된 레시피로 베이커리 제품을 만들 때 고농축 설탕에 대한 효모의 저항성이 중요합니다(내당성)
폭기 및 교반의 강도가 효모의 성장 속도에 미치는 영향.
효모를 재배할 때 영양 배지의 폭기가 필요하며, 이는 탄소 함유 영양 배지 1g당 0.8g O 2 로 정량적으로 표현됩니다. 폭기 강도를 계산하는 과정은 복잡하고 별도의 연구가 필요합니다.
효모 효소
제빵용 효모의 산업적 생산은 원칙적으로 당밀에서 수행되며, 당밀의 주성분은 자당입니다. 이와 관련하여, 효모 세포는 환경으로 쉽게 방출되는 엑소엔자임 β-프룩토푸라노시다아제를 능동적으로 유도한다. 이 효소는 항상 세포에 존재하며 세포막 외부에 집중되어 있습니다. 이와 관련하여 자당의 가수 분해는 효모 세포에 들어가기 전에 발생하며 효소 활성이 높으며 반제품 발효의 첫 번째 분부터 나타납니다.
효모가 자라는 영양 혼합물은 말토스를 함유하지 않아 효소 α-글루코시다아제(maltase)의 유도가 약하다. 엔도엔자임 α-글루코시다아제는 효모 세포의 세포질에 국한되어 있습니다. 맥아당이 발효되는 동안 탄수화물은 세포 속으로 침투하여 α-글루코시다아제라는 효소에 의해 두 개의 포도당 분자로 나뉩니다.
반죽을 느슨하게 하는 제빵 효모의 능력은 세포의 자이마제 복합체의 활성과 발효 가능한 당의 존재 여부에 달려 있습니다. 베이커리 생산의 밀가루 반제품의 설탕에는 자체 밀가루 설탕과 같은 여러 출처가 있습니다. 밀가루와 효모의 효소 작용에 의해 얻어지는 당류; 레시피에 따라 반제품에 첨가되는 설탕.
밀가루 자체 설탕의 양이 부족하여 기술 가치가 낮습니다. 탄소원으로 반제품 발효의 초기 단계에만 충분합니다. 반제품이 숙성되는 동안 설탕의 공급원은 전분으로, 밀가루의 전분 분해 효소의 작용으로 다음과 같이 분해됩니다. α -β -덱스트린과 맥아당. 처방 설탕을 포함하지 않는 베이커리 생산 반제품의 주요 "기술 설탕"은 맥아당입니다.
자당이 포함되지 않은 반제품에 압축 효모를 사용할 때 설탕 발효의 역학이 그림에 나와 있습니다.
쌀. 압착 효모를 이용한 반죽 발효 중 각종 당류의 발효 역학
반죽의 발효 중에는 설탕의 동시 발효가 거의 없습니다. 발효 초기에는 효모 세포가 포도당을 발효시키며, 과당과 맥아당의 발효는 각각 1시간과 2시간 후에 일어난다. 겨울 단지
효모 효소의 자이마제 복합체는 단당류를 알코올과 이산화탄소로 전환시킵니다. 포도당은 직접 발효되고, 과당은 이성질화 후 유도효소인 효모과당이성화효소(yeast fructoisomerase)에 의해 포도당으로 변합니다. 포도당과 자당을 발효시키는 효소는 구성 성분입니다. 자당은 다음의 작용에 따라 포도당과 과당으로 미리 전환됩니다. β -fructofuranosidase 효모와 그 역전율이 매우 높습니다.
배지에 말토스가 존재하면 효모 세포는 말토스를 세포로 운반하는 효소 maltopermease를 분비하고 효소 α -글루코시다아제(말타아제): 말토오스를 2개의 포도당 분자로 분할한 다음 이산화탄소와 에탄올의 형성과 함께 효소 복합체의 참여와 함께 효모에 의해 직접 발효됩니다. 맥아당의 발효에 관여하는 효소(maltpermease와 α -glucosidase)는 효모 세포가 이 이당류를 포함하는 배지에 들어간 후에만 형성됩니다. 그들은 유도성(적응) 효소입니다.
포도당 발효에서 과당 및 맥아당 발효로 효모를 전환하려면 효소 유도와 관련된 특정 기간이 필요하므로 이 기간 동안 반제품에서 가스 형성 속도가 약간 감소합니다. 맥아당 발효에 적응한 후 반죽에서 가스 형성 속도는 배지에 맥아당이 부족할 때까지 다시 증가합니다(그림).

쌀. 반죽 준비의 스펀지 방법 중 반제품에서 압축 효모의 가스 형성 속도의 역학
효소 maltopermease는 액체 모자이크 구조를 갖는 효모 세포의 세포질 막에 위치하며 지질 의존성 효소입니다. 효모의 세포질막에 위치한 효소계의 활성과 그 미세점도 사이에는 기능적 관계가 있는 것으로 알려져 있다. 따라서 투과 효소의 활성과 결과적으로 세포 내부의 효소 변형 강도는 막의 미세 점도에 따라 달라지며, 그 효과는 생화학적 발효 과정의 속도를 조절하는 데 사용될 수 있습니다.
유도 효모 효소의 분비는 배지에 축적되는 기질(말토오스)에 의존하기 때문에, 세포가 말토오스 배지에 적응하는 과정은 다소 길며 이는 아마도 반제품의 발효 기간에 영향을 미칠 수 있습니다. 밀가루 전분의 당화 과정을 가속화하기 위해 아밀로 분해 효소 제제가 반제품에 첨가되어 반죽의 발효성 설탕 함량이 증가하고 숙성 강화에 기여합니다.
밀가루 반제품에서 전분의 높은 당화 능력은 베이킹에 사용되는 효모의 유전적 특성을 변화시켜, 즉 특정 효모 효소의 생합성 및 분비를 조절함으로써 달성할 수 있습니다.
반제품 밀 제품의 알코올 발효에 대한 일반화된 모델이 그림 1에 나와 있습니다. 아홉.
빵 생산에서 효모의 역할을 특징으로하는 제시된 계획은 반제품의 효과가 생화학 적 변형의 전체 복합체에 달려 있음을 나타냅니다.

새로운 효모 균주의 선택을 위한 효과적인 방법으로서의 혼성화
잡종화는 기아, 주변 온도 감소 등과 같은 불리한 조건에서 포자를 형성하여 유성 재생산하는 효모의 능력을 기반으로합니다. 여러 산업의 정상적인 조건에서 효모는 발아에 의해 영양적으로 번식합니다. 교잡의 결과로 형성된 잡종은 원래의 부모 인종보다 번식 에너지가 증가했습니다.
잡종 saccharomycetes가 생산을 위해 선택되는 징후 :
크기가 7x11 마이크론 이상인 대형 셀;
말타아제 활성은 분 이하입니다.
45분 이하의 리프팅 힘;
당밀 저항 100%
효모 경주
현재, 일반적으로 효모 생산에서 다음 유형의 효모 균류를 사용하는 것이 허용됩니다. 사카로마이세스 세레비지애다른 인종. 인종은 주어진 종의 모든 주요 특성을 유지하면서 생산 특성을 특징짓는 이차적이지만 지속적인 특성으로 구별되는 다양한 미생물로 이해됩니다. 매우 자주 인종을 균주라고 하는데, 이는 잘못된 균주이기도 하며 실험실 조건에서만 테스트된 이 종의 변종이기 때문입니다(Semikhatova N.M., 1980). 인종이나 혈통으로같은 종 내에서 이차적 특성이 서로 다른 별개의 미생물이라고 합니다. 동시에, 종족은 지속적인 2차 형질을 가지며, 균주는 불안정하며 새로운 배지에서 자랄 때 손실될 수 있습니다(Matveeva I.V., Belyavskaya I.G., 2001).
효모 생산 및 베이커리 공정의 개선과 관련하여 효모에 대한 새로운 요구 사항이 점점 더 많아지고 있습니다. 활발한 생산 인종을 특징짓는 특성 선택에 대한 견해도 변화하고 있습니다. 이전에는 주로 문화 및 형태학적 특성에 따라 선별이 이루어졌으나, 이제는 효모의 생화학적 특성과 효소적 특성에 기반을 두고 있습니다.
효모의 생산 배양은 반제품의 장기간 준비, 높은 효소 활성을 포함하는 빵 준비의 다단계 기술 모드에서 특히 중요한 높은 비성장률을 가져야 합니다.
베이킹에 사용되는 개별 효모 균주의 형태학적 및 물리화학적 특성과 기술적 매개변수의 특성은 다음과 같습니다.
1939년에 원래 철회됨 레이스 톰스카야 7 E.A. 플레바코와 N.G. 톰스크 효모 공장의 압축 효모로 만든 마카로바. 이 인종은 성장 물질, 특히 비타민을 요구하는 당밀 배지 구성에 대한 저항성이 특징입니다. 이 품종에서 얻은 압축 효모는 저장 중에 안정적이며 높은 β -fructofuranosidase 활성, 그러나 약함 α -글루코시다아제 활성(말타아제 활성 160이상 분).
레이스 오데사 14 1958년 오데사 효모 공장 3.I.에서 분리되었습니다. 수입 건조 효모 샘플의 Vishnevskaya. 이 문화는 높은 생산 활동이 특징입니다. 누룩건조에 강하고 눌렀을 때 보관 중에 안정적입니다. 말타아제 활성은 95 분,자이마제 - 45 분배양은 영양 배지의 구성, 특히 성장 물질을 요구합니다. 그러나 높은 수율과 효소 활성으로 인해 산업계에서 널리 사용됩니다.
균주 L-441 LO GosNIIKhP에서 Odessa 14 인종 효모의 자연적 다양성에 기반한 선택에 의해 사육됩니다. 균주 L-441은 높은 생산성, 라피노스 발효, 유해한 불순물 및 병원성 미생물에 대한 저항성, 높은 비성장률 및 우수한 특성을 특징으로 합니다. 상업용 베이커리 효모: 리프팅 포스 44 -45 분,말타아제 활성 92-95 분, 96 시간 동안 35 ° C의 온도에서 저항.
균주 I-1 14종 효모의 순수한 산업 배양에서 표적 선택에 의해 양이율 효모 공장에서 자란다. 이 균주는 수년 동안 생산 조건에서 테스트되었습니다. 문화는 높은 생산 활동과 높은 성장 온도 (37-38 ° C)에 대한 저항성을 가지며 이는 국가의 남부 지역에 위치한 식물에 매우 중요합니다. 상업용 효모의 리프팅 포스 - 40-47 분,자이마제 활성 32-44 분
레이스 키예프스카야 21 1960년 M.K. 수입 건조 효모의 리드만(Reidman)은 생체 자극제로 다중 활성화하는 방법을 사용합니다. 배양물은 성장 물질에 대한 요구가 없고 건조에 잘 견디며 좋은 자이마제(60 분)및 밀타제(100 분)활동.
하이브리드 레이스 176, 196-6 및 262산업용 효모에 대한 기본 요구 사항을 충족하고 산업용으로 권장됨: 말타제 활성 65-75 분,자이마제 42-57 분,높은 성장률.
사육 새로운 균주 739, 743, 608, 616, 722높은 효소 활성으로. 압축 및 건조 효모의 생산에 사용되는 균주 LV-7이 개발되었습니다. 이 균주는 효모 생산을 감염시키는 당밀 및 미생물총의 불순물에 대한 내성 증가가 특징이며, 생산성 증가가 특징이며 트레할로스 농도 측면에서 대응 물보다 2배 높습니다. 압축 효모 균주 LV-7의 리프팅 힘은 43-47입니다. 분,삼투 감도 - 6-10 분.
효모 균주 616건조 효모 생산에 사용되며 효모 효소 시스템의 활성에서 인종 14를 능가합니다. 효모의 말타아제 활성은 67입니다. 분,자이마제 - 55 분
균주 722좋은 말타아제 (54 분),자이마제(43 분)활동, 리프팅 포스(46 분)및 삼투 감도(5-10 분).
균주 739높은 생산성, 증가된 효소 활성이 특징입니다. 효모는 포도당, 과당, 자당, 말토오스, 라피노오스, 갈락토오스를 완전히 발효시킵니다. 효모의 자이마아제, 말타아제 활성 및 리프팅력은 각각 54, 61 및 56입니다. 분.
효모 균주 사카로마이세스 세레비지애 39/15는 발효 활성이 좋으며 이를 사용하면 반죽의 발효 기간을 35까지 줄일 수 있습니다. 분
건조 효모 생산을 위해 균주가 사용됩니다. 사카로마이세스 세레비지애 93, 높은 생산성으로 효소의 활성 복합체. 겨울 활동은 45입니다 분,말타아제 - 53 분,리프팅 포스 - 45 분
XII 인종과 균주를 교배하여 얻은 잡종 균주 512 사카로마이세스 디아스태쿠스 , 3배체이며 비타민 D(에르고스테롤) - 2.8의 합성 증가가 특징입니다. B 1 - 34; B 2 - 20; B 6 46, PP - 36(㎍/세포). zymase, maltase 활성 및 삼투 감도 지수는 70, 200 및 14입니다. 분각기.
균주 5 발효에 사용되는 Yablochny-3 효모 균주의 세포를 교배하여 얻은 사과 주스및 건조된 빵 효모의 생산에 사용되는 균주 722. 이 균주의 특징은 높은 발효 활성입니다. zymase, maltase 활성 및 삼투압 지수는 85, 95 및 15입니다. 분
균주 69는 건조된 프랑스 효모로부터 분리된 Jambul-60 효모 레이스와 균주 10을 교배하여 얻었다. 균주 69는 각각 높은 성장률, 자이마제 및 말타제 활성을 갖는다 45 분그리고 80 분,고온 (40-45 ° C)에 대한 내성.
속 다른 종의 대표 사카로마이세스 효모이다 사카로마이세스 미성년자 , 호밀 누룩에서 발견. 이 작은 원형 또는 약간 타원형의 효모는 1872년 Engel에 의해 처음으로 분리되고 기술되었습니다. 그들은 포도당, 과당, 자당, 갈락토스, 라피노스를 발효 및 동화시키고, 유당, 자일로스, 아라비노스, 글리세린을 발효 및 동화시키지 않으며, 전분과 섬유를 분해하지 않습니다. 이 종의 특징은 맥아당과 단순 덱스트린을 발효하거나 동화시키지 않는다는 것입니다. 최적의 온도는 25-28 ° C이며 온도를 35 ° C로 올리면 우울한 효과가 있습니다. 누룩 사카로마이세스 미성년자 더 큰 내산성(산도 14-16 ° 및 pH 3.0-3.5에서 잘 발달함)과 내알코올성으로 구별됩니다. 사카로마이세스 세레비지애 .
현재, 유도된 돌연변이 유발, 교잡, 적응과 같은 현대적인 방법을 사용하여 새로운 효모 균주의 육종에 대한 작업이 계속되고 있습니다. 이것은 베이커리 제품 준비를 위한 현대 기술의 구현에 필요한 고정된 품질 특성을 가진 순수한 미생물 배양의 효과적인 선택에 기여합니다.
제빵 효모의 종류
베이커리 제품의 준비에는 베이커리 프레스, 건조, 인스턴트 효모, 효모 우유 및 액체 발효 효모가 사용됩니다.
압축 효모 – 기술적으로 순수한 효모 배양물입니다. 사카로마이세스 세레비수이 , 61-75%의 습도로 연탄으로 형성됩니다. 배양액은 압축 또는 진공 청소기로 상업용 효모가 얻어질 때까지 배지를 집중적으로 통기하는 조건에서 자궁 및 종자 효모의 바이오매스를 축적함으로써 특수 영양 배지에서 성장합니다. 압축 효모 1g에는 100~150억 개의 세포가 들어 있습니다.
건조 효모 – 일정한 조건에서 수분함유량 8~10%로 건조시킨 압축효모로 예비보습후 사용합니다.
인스턴트 효모 – 현대적인 재배 조건, 건조 방법 및 보호 첨가제 및/또는 유화제를 사용하여 특정 사카로마이세테스 균주를 기반으로 제조된 반죽에 첨가하기 전에 재수화를 필요로 하지 않는 고활성 건조 효모.
효모 우유 (분리 효모 ) – 400-450g / l 농도의 효모 현탁액으로 분리 후 얻어 압축 효모 대신 사용합니다.
액체 효모 – 제과점에서 특별히 제조된 당화 양조주를 기본으로 하여 호열성 유산균으로 발효시킨 후 그 위에 효모를 배양한 반제품 사카로마이세스 . 액체 효모는 생물학적 게스트 팽창제로 사용되거나 빵의 품질을 향상시키는 수단으로 사용됩니다. 액체 효모 1ml에는 7천만~1억 2천만 개의 세포가 들어 있습니다.
현대 베이커리 생산 기술의 발전에는 특정 기술 계획에 맞게 조정된 효모의 사용이 필요하므로 많은 기업과 회사에서 삼투압성, 반건조 냉동 효모, 추위에 민감하고 프로피온산칼슘에 내성이 있는 효모를 생산합니다. 베이커리 제품을 위한 기성품 혼합물에 사용.
삼투성 누룩 밀가루 중량의 10% 이상인 과립 설탕 함량으로 반죽을 준비하기 위한 것입니다. 삼투압 효모의 특징은 인버타제 함량이 낮고 트레할로스와 글리세롤을 합성하는 능력으로 삼투압을 낮추고 세포 내 수분 손실을 보상할 수 있습니다.
반건조 냉동 효모 베이커리 및 패스트리 제품용 급속 냉동 반죽 반제품 기술에 사용하기 위한 것입니다. 건조 물질 함량은 75-77%입니다. 효모 생산 중 건조 후 동결되어 보관 중 안정성이 높아집니다. 반건식 냉동 효모의 특징은 발효 과정의 시작이 지연되고 저온 저장 동안 냉동 반죽에서 특성의 안정성이 지연된다는 것입니다.
저온에 민감한 효모 , 4-12 ° C의 온도 범위에서 극도로 낮은 효소 활성과 30-40 ° C의 온도에서 표준 활성이 특징입니다. 이를 통해 소매 반죽 준비에 사용할 수 있습니다. 이 효모로 준비한 반죽 조각은 발효 과정에 따른 변화 없이 3-7°C의 온도에서 며칠 동안 보관할 수 있으므로 신속하게 동결할 필요가 없습니다.
칼슘 프로피오네이트 저항성 효모 , 베이커리 제품에서 감자 질병을 예방하기 위한 수단으로 프로피온산 칼슘을 첨가하여 제조한 반죽에 대한 내산성 및 적응성이 증가하는 것이 특징입니다.
기성품 혼합물 생산용 효모 (프리믹스 ) , 산소와 수분이 있는 상태에서 보관할 수 있으며 예비 수화도 필요하지 않습니다. 효모는 보호 과립의 특수 구조로 인해 이러한 특성을 가지고 있습니다. 이 보호 과립은 특수한 껍질을 가지고 있고 구조의 높은 다공성을 특징으로 하며, 이는 빵집 생산의 과립 및 반제품의 빠른 용해에 기여합니다.
비활성화 효모 – 발효 능력은 없으나 효소 활성을 갖는 제품. 이 효모는 신축성과 신축성이 필요한 반죽을 위한 천연 복원 개선제입니다.
다양한 유형의 효모 사용의 효과는 영양 배지의 구성에 따라 설탕 발효의 기본 운동 법칙, 환경 매개 변수의 영향, 효모 대사의 특성에 대한 지식에 의해 결정되며 다음과 같이 결정됩니다. 효모의 생리적, 생물학적 및 기술적 특성.
압축 효모는 밀 반죽과 호밀과 호밀의 혼합물로 반죽을 만드는 데 사용됩니다. 밀가루레시피, 생산 방법 및 공정 매개변수에 따라 0.1~8중량%의 밀가루.
압축 효모는 1:2에서 1:4의 비율로 물에 미리 준비된 효모 현탁액 형태로 반제품에 도입됩니다. 건조 효모의 사용에는 재수화의 예비 단계, 때로는 활성화가 포함됩니다. 인스턴트 효모의 경우 예비 준비가 필요하지 않으며 느슨한 형태로 반죽에 도입됩니다. 다양한 종류의 제빵 효모를 사용할 때의 비교 특성이 표에 나와 있습니다. 1.
반죽의 효모 양과 활성에 따라 달라지는 중요한 요소는 반제품의 발효 기간 및 온도와 같은 기술 과정의 매개 변수입니다. 반죽의 발효 과정이 감소함에 따라 효모의 양이 증가합니다. 발효 온도 계수 값과 발효 온도 사이에 직접적인 패턴이 나타났습니다. 온도가 25°C에서 35°C로 증가하면 발효 강도가 약 2배 증가합니다.
효모의 복용량은 반죽 준비 방법에 따라 다르며, 결정 매개 변수는 공정 기간입니다. 빵집 생산의 경우 반죽 준비 방법에 따라 다음과 같은 양의 압축 효모가 사용됩니다. 스펀지 방법 - 0.5-1.0%; 짝을 이루지 않은 방법 - 2.0-2.5%; 단상 가속 방법 - 밀가루의 3.0-6.0 중량%.
실제적으로 중요한 것은 반죽 준비 방법 및 결과적으로 반제품의 발효 기간에 따라 다양한 당과 관련하여 효모의 발효 활성을 구별하는 것입니다. 스펀지 및 짝이 없는 방법의 경우(총 숙성 시간은 210-350입니다. 분)효모의 높은 말타제 활성은 최적의 반죽 특성과 우수한 빵 품질을 달성하는 데 필수적입니다. 반죽의 발효 과정에서 발효 시간은 180-240입니다. 분,혐기성 맥아당 가루 배지에 대한 효모 세포의 적응이 일어나므로 반죽에서 가스 형성의 강도는 짝을 이루지 않은 방법과 비교하여 효모의 초기 말타제 활성에 훨씬 덜 의존합니다.
대량의 반죽발효를 배제하고 총숙성시간이 70~100 반제품인 가속기술 구현시 분,유도 α - 일반적으로 70-90에서 시작하는 효모에 의한 글루코시다아제 분반죽 발효 과정의 시작부터 기술 과정의 과정에 큰 영향을 미칠 수 없습니다. 따라서 효모의 높은 자이마제 활성이 우선적으로 중요합니다. 베이커리 제품 준비에 가속 기술을 사용할 때 권장 사항은 반죽에 최소 2%의 과립 설탕을 추가하는 것입니다.
반죽의 효모 양에 영향을 미치는 중요한 요소는 조리법, 즉 설탕과 지방 제품의 양입니다. 반죽에 설탕과 지방 함유 제품이 있으면 효모의 효소 활성과 결과적으로 그 양에 영향을 미칩니다. 반죽의 밀가루 질량에 7 % 이상의 과립 설탕을 첨가하면 효모 세포의 plasmolysis 과정이 시작되어 중요한 활동이 감소합니다. 반죽에 지방 제품을 5% 이상 첨가하면 효모 세포 표면의 지방 흡착으로 인해 가스 형성이 감소하여 용해성 영양소가 세포막을 통과하는 속도가 느려지거나 중단됩니다. , 효모 대사 과정을 방해합니다. 이것이 페이스트리 제품의 반죽에서 효모의 양을 밀가루의 4-6 중량%까지 늘리거나 설탕과 설탕의 도입을 제공하는 반죽 채우기 단계를 기술 공정에 도입하는 것이 좋습니다. 반죽 발효의 마지막 단계에서 지방 제품.
효모의 유형 및 최적 복용량 선택, 반제품 베이커리 제품의 발효 기간은 발효 중에 발생하는 법칙, 다양한 유형의 효모의 생명 공학 특성에 대한 지식, 레시피 구성 요소의 영향 메커니즘을 기반으로합니다. 기술 과정의 매개 변수, 반죽 준비 방법과 관련하여.
맥주 효모의 인종
양조는 비교적 낮은 온도에 적응된 하면 발효 효모를 사용합니다. 맥주효모는 미생물학적으로 순수해야 하며 응집하는 능력이 있어야 하며 발효 장치의 바닥에 빠르게 가라앉고 특정 맛과 향을 지닌 맑은 음료를 제공해야 합니다. 하면 발효 Froberg(Saccharomyces cere-visiae Froberg)의 맥주 효모, V 및 776 인종의 효모는 강하게 발효되어 플레이크를 쉽게 생성합니다.
양조장에서는 20 세기 초에 자란 776 품종의 효모가 널리 보급되었습니다. 이 효모는 발아율이 낮은 보리를 발아시켜 얻은 맥아 또는 맥아를 발아하지 않은 재료를 첨가하여 만든 맥아즙을 발효시키는 데 특히 유용한 것으로 간주됩니다. 효모 레이스 776은 적당히 발효 중이며 11 % 농도의 맥아 즙에 대한 주요 발효 중에 약 2.7 % СО 2를 형성합니다. 세포는 난형이며 길이 8-10 미크론, 너비 5-6 미크론입니다. 효모 질량의 증가는 1:5.4입니다. 라이트닝 능력은 만족스럽습니다.
다른 효모 중에서 양조장은 발효 에너지, 침전 능력 및 성장 에너지가 다른 인종 11, 41, 44, S-Lvovskaya 등을 사용합니다.
Race 11 효모 - 비옥도가 높고 정화 능력이 우수합니다. 레이스 11 효모로 만든 맥주는 맛이 좋습니다. 이 경주는 양조장에서 널리 퍼졌습니다.
Race 41 효모 - 적당히 발효되고 강수 경향이 좋습니다. 레이스 41로 맥아즙을 발효시키면 깔끔한 맛의 부드러운 맥주가 나옵니다.
Race 44 효모 - 적당히 발효. 정착 능력이 좋습니다. 그들은 맥주에 완전한 풍미를 제공하고 경도가 증가된 물 생산에 사용될 때 좋은 결과를 제공합니다.
Race S 효모 - 적당히 발효. 정착 능력이 좋습니다. 그들은 순하고 깨끗한 맛의 맥주를 제공합니다.
Race P 효모는 적당히 발효되어 맥주를 잘 빛나게 하고 쾌적하고 깨끗한 맛을 제공합니다.
Race F 효모는 우수한 정화 능력이 특징이며 맥주에 기분 좋은 향을 부여합니다. 인종은 외래 미생물의 작용에 저항합니다.
Race A 효모(Riga 양조장 "Aldaris"에서 분리)는 7-8일 안에 맥아즙을 발효시키고 맥주를 잘 정화하며 감염에 강합니다.
원래 인종 11의 효모보다 훨씬 더 높은 발효 에너지를 가진 많은 고도 발효 효모 균주(28, 48, 102)가 맥주 및 음료 산업의 전 러시아 연구소(28, 102)에서 다양한 육종 방법에 의해 얻어졌습니다. 48, 102).
상면 발효 맥주 효모는 영국에서 포터를 준비하는 데 널리 사용됩니다. 그들은 또한 베를린 라이트 맥주 및 기타 음료를 준비하는 데 사용됩니다. Velvet 맥주의 제조를 위해 단당류와 맥아당을 집중적으로 발효시키지만 자당, 라피노스 및 유당은 발효하지 않는 균주 191K가 사용됩니다.
^ 와인 효모 레이스
포도주 양조법은 빠르게 증식하고 다른 유형의 효모와 미생물을 억제하는 능력이 있으며 포도주에 적절한 부케를 주는 효모를 높이 평가합니다. 포도주 양조에 사용되는 효모는 Saccharomyces ellipsoideus의 일종에 속합니다. 그들의 세포는 직사각형 타원형입니다. 효모는 포도당, 과당, 자당, 맥아당을 격렬하게 발효시킵니다. 다른 지역과 다양한 젊은 와인에서 이 종의 여러 다른 품종이나 인종이 서로 구별됩니다. 포도주 양조에서 거의 모든 효모 생산 문화는 고유한 지역 기원입니다. 여기에는 Magarach 7, Massandra 3, Pino 14, Kakhuri 및 기타 여러 경주가 포함됩니다. 이러한 인종과 함께 1892년과 1893년에 독일에서 분리된 슈타인베르그(Steinberg) 경주와 샴페인-오(Champagne-Au) 경주와 같은 일부 외국 경주도 사용됩니다.
와인 효모의 대부분은 하면 발효 효모에 속합니다.
화이트 테이블 와인을 준비하기 위해 Pinot 14, Feodosia 1/19, Aligote, Riesling Anapsky가 사용됩니다.
Pinot 14 품종은 난형 세포를 가지고 있으며 11.57%의 알코올을 형성하며 20%의 설탕 함량으로 포도 머스트를 잘 발효시킵니다. 발달 및 발효를 위한 최적 온도는 18: -25 ° C입니다. 이 종족은 추위에 강하고 산성에 강합니다. 최적의 pH 값은 2.9-3.9입니다.
Race Theodosia 1/19 - 세포가 크고 먼지가 많으며 에너지가 넘치며 포도 머스트를 빠르게 발효시키고 잘 발효시킵니다. 넓은 발효 온도 범위(9~35°C)를 가지며 내한 및 내열로 사용할 수 있습니다.
알리고테 효모에는 여러 종족이 있으며 모두 강력하고 발효 에너지가 높습니다. Riesling Anapsky 효모는 또한 활발하게 발효되는 효모입니다.
강한 와인을 준비하기 위해 Massandra 3 레이스는 먼지가 많은 달걀 모양의 셀과 함께 사용됩니다. 최적의 pH 값은 3.7-4.05입니다. 최적의 발효 온도는 18-20 ° C입니다. 설탕 함량이 20%인 포도는 완전히 발효됩니다. 발효된 농축포도(설탕 30%)는 부피로 11.8%의 알코올을 형성하고 발효되지 않은 8.7%의 설탕을 남깁니다.
Magarach Institute의 첫 포도 재배 125주년을 기념하여 명명된 Race Magarach 125는 스트롱 와인과 디저트 와인을 생산하는 데 사용됩니다. 당도 27~30%, 내한성으로 고농축 포도 머스트를 잘 발효시키는 품종입니다.
Race Kakhuri 2는 샴페인 와인 재료 및 와인 준비에 널리 사용됩니다. 20%의 설탕 함량으로 포도 머스트를 발효시켜 부피 기준으로 11.4%의 알코올을 형성하고 0.28%의 설탕은 발효되지 않은 상태로 유지합니다. 이 경주는 매우 내한성이 있으며 (14-15 ° C의 온도에서 2 일째에 발효됩니다) 잘 발효됩니다. 최적의 pH 값은 3.4-3.6입니다.
병에 담긴 와인을 샴페인화하는 데 사용되는 Champagne 7 종족은 Kakhuri 5 종족과 분리되어 교반하기 어려운 침전물이 형성되는 것이 특징입니다. 4-9 ° C의 온도에서 집중적으로 발효되지만 wort는 5-6 일에만 발효됩니다.
와인 효모 중에서 레닌그라드(Leningrad) 레이스가 가장 내한성이 강한 것으로 간주되며, 아쉬하바트(Ashgabat) 레이스가 가장 내열성이 높은 것으로 간주됩니다.
셰리 생산에는 Saccharomyces oviformis 종의 일종인 특별한 효모 레이스가 사용됩니다. 셰리 효모는 불완전한 배럴에서 와인 표면에 필름을 형성하여 와인이 특별한 꽃다발과 맛을 얻도록 발달합니다.
가장 중요한 생산 특성에 따라 신중한 선택을 통해 높은 필름 형성 능력을 가진 여러 품종의 셰리 효모(13, 15 및 20)를 선택했습니다. 나중에 Jerez 20 레이스를 사용한 생산에서 더 효율적인 레이스 Jerez 20-C가 선택되어 많은 셰리 공장에서 널리 사용되었습니다.
과일 및 베리 포도주 양조에서는 다양한 과일 및 베리 주스에서 분리된 선택된 효모 인종이 사용됩니다. 과일 및 베리 주스는 생산에 필요한 모든 특성을 지닌 효모가 풍부하고 원래 과일 및 베리 주스의 발달 조건에 생물학적으로 적응합니다. 따라서 딸기 주스에서 분리한 효모 균주는 딸기 주스를 발효시키는 데 사용하고, 체리 주스에서 분리한 효모 균주는 체리 주스 등을 발효하는 데 사용합니다.
과일 및 베리 포도주 양조법에 다음 계통이 널리 퍼져 있습니다. 사과 46, 58, 크랜베리 17, 건포도 16, 링곤베리 3, 7, 10, 라즈베리 7/5, 25, 28, 28/10, 체리 3, 6, 딸기 7 , 4, 9.
명명 된 효모 균주는 정상적인 발효 과정, 발효의 완전성, 빠른 정화 및 와인의 좋은 맛을 보장합니다. 그들은 포도당, 과당, 자당, 말토오스, 갈락토오스를 발효하고 유당과 만니톨을 발효시키지 않습니다.
그들은 효모 레이스 Moscow 30, Yablochnaya 7, Vishnevaya 33, Chernoscurorodinovaya 7, Raspberry 10 및 Plum 21의 과일 및 베리 포도주 양조에 성공적으로 사용되었습니다. 크랜베리 맥아즙의 발효에는 효모 Moscow 30의 순수 배양이 권장됩니다. Apple 7 및 Cherry 33 - 사과 맥아즙 발효용; 블랙커런트 7 및 체리 33 - 블랙커런트와 체리 맥아즙 발효용.
^
4 알코올 발효의 화학. 알코올 발효의 2차 및 부산물
알코올 발효는 일련의 효소적 과정이며, 최종 결과는 6탄당 분해로 알코올과 CO 2가 형성되고 효모 세포에 에너지가 전달되며 이는 생명 유지에 사용되는 새로운 물질의 형성에 필요합니다. 성장과 번식을 포함한 과정. 화학적 특성으로 인해 알코올 발효는 생물학적 촉매 인 효소의 작용으로 발생하는 촉매 과정입니다.
알코올 발효에 대한 현대 이론은 세계 여러 나라의 많은 과학자들의 연구 결과입니다.
Lebedev, Kostychev, Favorsky, Ivanov, Engelhardt와 같은 뛰어난 러시아 과학자들의 작업은 발효 과정을 설명하는 데 매우 중요했습니다.
현대 개념에 따르면 알코올 발효는 12개의 중간 생성물을 형성하는 다양한 효소에 의해 촉매되는 당 분해의 복잡하고 연속적인 과정입니다.
1 포도당 전환의 초기 단계는 글루코시나아제 효소의 참여와 인산화 반응입니다. 효모 세포에서 발견되는 ATP 분자의 인산 잔기가 포도당 분자에 부착되어 포도당-6-인산이 형성되고 ATP가 ADP로 전환됩니다.
C 6 H 12 O 6 + ATP → CH 2 O (H 2 PO 3) (CHOH) 4 CHO + ADP
포도당 포도당-6-인산염
ATP 분자의 인산염 잔기를 포도당에 추가한 결과 후자의 반응성이 증가합니다.
2 포도당 6-인산은 효소 포도당 인산염 이성화효소의 작용하에 이성질화되어 과당의 형태로 전환됩니다.
CH 2 O (H 2 PO 3) (CHOH) 4 CHO → CH 2 O (H 2 PO 3) (CHOH) 3 COCH 2 OH
포도당-6-인산과당-6-인산
3 또한, 효소 phosphofructokinase의 작용으로 두 번째 ATP 분자에서 fructose-6-phosphate와 fructose-1,6-diphosphate로 또 다른 인산 잔기가 이동하고 새로운 ADP 분자가 형성됩니다.
СН 2 О (Н 2 РО 3) (СНОН) 3СОСН 2 ОН + ATP →
과당-6-인산
→ СН 2 О (Н 2 РО 3) (СНОН) 3 СОСН 2 О (Н 2 РО) + ADP
과당-1,6-이인산
에스테르 글루코스-6-포스페이트 및 프럭토스-6-포스페이트는 Emden 에테르라고 하는 평형 혼합물을 형성하고 70-75%의 로비슨 에테르(포도당)와 25%의 노이버그 에테르(과당)로 구성됩니다.
준비 과정은 과당-1,6-이인산의 형성으로 끝납니다. 고 에너지 인산염 결합이 이동하고 6탄당이 불안정한 옥시 형태로 변형되는 알코올 발효 단계로, 이는 추가 효소 변형에 쉽게 노출됩니다.
4 다음으로 가장 중요한 단계는 해체입니다 - 과당 이인산의 탄소 사슬을 끊고 두 개의
인산화 분자. 과당 분자의 말단에 있는 인산 잔기의 대칭 배열은 중간에 있는 탄소 사슬의 절단을 촉진합니다. 과당 이인산은 포스포글리세롤 알데히드와 포스포디옥시아세톤의 두 가지 삼당으로 분해됩니다. 반응은 효소 알돌라제에 의해 촉매되며 가역적입니다.
СН 2 О (Н 2 РО 3) (СНОН) 3 СОСН 2 О (Н 2 РО) → CH 2 О (Н 2 Р0 3) СОСН 2 ОН +
과당-1,6-이인산 포스포디옥시아세톤
채널 2 0 (H 2 ROS) 채널 (4)
3-포스포글리세롤 알데히드
알코올 발효 중 추가 변형의 주요 역할은 3-포스포글리세롤 알데히드에 속하지만 발효 액체에서는 미미한 양으로만 발견됩니다. 이것은 효소 triose phosphate isomerase(5.3.1.1)의 작용하에 케토스 이성질체가 알도스 이성질체로 또는 그 반대로 상호 전이하기 때문입니다.
СН 2 0 (Н 2 Р0 3) СОСН 2 ОН, £ СН 2 0 (Н 2 Р0 3) СНОНСНО
포스포디옥시아세톤 3-포스포글리세롤 알데히드
phosphoglycerol al-dehyde의 추가 전환으로 phosphodioxyacetone의 이성질체화 과정에서 새로운 양이 형성됩니다.
5. 다음 단계는 3-포스포글리세롤 알데히드 두 분자의 산화입니다. 이 반응은 조효소가 NAD(니코틴아미드-아데닌-디뉴클레오티드)인 트리오스 포스페이트 탈수소효소(1.2.1.12)에 의해 촉매됩니다. 매체의 인산은 산화에 관여합니다. 반응은 다음 방정식에 따라 진행됩니다. 2CH 2 O (H 2 PO 3) CHONCHO + 2H 3 PO 4 + 2NAD Triose phosphate dehydrogenase ->
3-포스포글리세롤 알데히드
-> - 2СН 2 0 (Н 2 Р0 3) SNONSOO w (H 2 P0 3) + 2NAD Н 2 (5)
1,3-디포스포글리세롤산
3-phosphoglycerol aldehyde 분자는 인산염을 붙이고 수소는 보효소 NAD로 옮겨져 환원된다. 3-포스포글리세롤 알데히드의 산화로 인해 방출된 에너지는 형성된 1,3-디포스포글리세롤의 고에너지 결합에 축적됩니다.
산.
6. 또한, 1,3-디포스포글리세린산의 인산 잔기
효소의 참여로 고에너지 결합을 포함하는 당신
포스포글리세르산 키나제(2.7.2.3)는 ADP 분자로 전달됩니다.
3-포스포글리세르산이 형성되고, ADP, 획득
추가 고에너지 연결, ATP로 전환:
2СН 2 0 (Н 2 Р0 3) СНОСОО с (Н 2 Р0 3) + 2ADP-> 2СН 2 0 (Н 2 Р0 3) CHOHCOOH +
1,3-디포스포글리세르산 3-포스포글리세르산
7. 그런 다음 효소 phosphoglyceromutase의 작용하에
(2.7.5.3) 인산 잔기는 세 번째에서 이동
탄소를 두 번째로, 결과적으로 3-포스포글리세롤산
lot은 2-phosphoglyceric acid로 변합니다.
2СН 2 (Н 2 Р0 3) CHOHCOOH ^ t 2CH 2 0HCH0 (Н 2 Р0 3) COOH. (7)
3-포스포글리세르산 2-포스포글리세르산
8. 다음 단계는 2-포스포릴의 탈인산화
포글리세린산. 이 경우 2-포스포글리세롤산
탈수에 의한 효소 enolase(4.2.1.11)의 작용하에 lot
tirovanie(물 손실)가 포스포에놀피루빅으로 변합니다.
우박산:
2CH 2 ONSNO (H 2 P0 3) COOH qt 2CH 3: CO co (H 2 P0 3) COOH + 2H 2 0. (8)
2-포스포글리세르산 코쇼에놀피루브산
이 변환 동안 분자 내 에너지의 재분배가 발생하고 대부분은 고에너지 인산염 결합에 축적됩니다.
9. 매우 불안정한 포스포에놀피루브산
쉽게 탈인산화, 나머지 인산으로
효소 pyruvate kinase (2.7.1.40)의 작용으로 전달됩니다
ADP 분자에 대한 고에너지 결합과 함께. 결과적으로
더 안정적인 케토 형태의 피루브산이 형성됩니다.
당신과 ADP는 ATP로 바뀝니다.
2CH 2: CO sy (H 2 R0 3) COOH + 2ADP - * 2CH 3 ^ COOH + 2ATF. (삼)
포스포에놀피루빅
산성산
10. 파이루브산(Pyruvic acid) 효소의 작용하에
루베이트 데카르복실라제(4.1.1.1)는 절단으로 탈카르복실화됩니다.
CO 2 형성 및 아세트알데히드 형성:
2CH 3 ^ COCOOH - * 2CO 2 + 2CH 3 CHO. (십)
피루브산 알데히드
11. 알코올 탈수소 효소의 참여로 아세트산 알데히드-
rogenase(1.1.1.1)는 형성된 NAD-H 2와 상호작용합니다.
이전에 phosphoglyceric aldehyde가 phospho-로 산화되는 동안
글리세린산[참조. 식 (5)]. 결과적으로 식초
알데히드는 에틸알코올로 환원되고, 조효소
NAD-H 2는 다시 재생됩니다(NAD로 산화됨):
2CH 3 CHO + 2 OVER H 2 지 2CH 3 CH 2 OH + 2 OVER. (열하나)
따라서 발효의 마지막 단계는 아세트알데히드를 에틸알코올로 환원시키는 반응입니다.
알코올 발효의 고려된 반응 주기에서 각 포도당 분자에서 2분자의 알코올과 2분자의 CO2가 형성됨을 알 수 있습니다.
알코올 발효 과정에서 4개의 ATP 분자가 형성됩니다[참조. 방정식 (6)과 (9)], 그러나 그 중 두 개는 육탄당의 인산화에 사용됩니다[참조. 방정식 (1) 및 (3)]. 따라서 ATP는 2g-mol만 저장됩니다.
ADP에서 ATP의 각 그램 분자를 형성하는 데 41.9kJ가 소비되고, 83.8kJ가 두 ATP 분자의 에너지로 각각 전달되는 것으로 이전에 표시되었습니다. 결과적으로 1g-mol의 포도당을 발효할 때 효모는 약 84kJ의 에너지를 얻습니다. 이것이 발효의 생물학적 의미입니다. 포도당이 CO2와 물로 완전히 분해되면 2874kJ가 방출되고 1g-mol의 포도당이 CO2와 H2O로 산화되면 호기성 호흡 과정에서 2508kJ가 축적됩니다. 잠재적 에너지. 따라서 에너지 관점에서 발효는 저비용 공정입니다.
개별 당의 발효는 효모 세포로의 확산 속도 때문에 특정 순서로 발생합니다. 포도당과 과당은 효모에 의해 가장 빨리 발효됩니다. 그러나 자당 자체는 발효 초기에도 맥아즙에서 사라집니다(역전). 효모 세포막의 β-fructofuranosidase(3.2.1.26)에 의해 가수분해되어 세포에서 쉽게 사용되는 육탄당(포도당 및 과당)을 형성합니다. 맥아즙에 과당과 포도당이 거의 남지 않으면 효모가 맥아당을 소비하기 시작합니다.
§ 5. 알코올 발효의 2차 및 부산물
알코올과 CO2를 제외하고 효모에 의한 당의 발효로 인한 모든 물질은 알코올 발효의 2차 산물입니다. 그 외에도 설탕이 아닌 발효성 기질에서 발견되는 다른 물질에서 생성되는 알코올 발효 부산물이 있습니다. 여기에는 아밀, 이소아밀, 이소부틸 및 퓨젤 오일로 알려진 기타 알코올이 포함됩니다.
알코올 발효의 이차 생성물 중에는 글리세린, 아세트알데히드, 피루브산, 아세트산, 숙신산, 시트르산 및 젖산, 아세토인(아세틸메틸-카르비놀), 2,3-부틸렌 글리콜 및 디아세틸이 알려져 있습니다. 호기성 조건에서 피루브산은 트리카르복실산 회로(크렙스 회로)의 출발 물질이기도 하며, 이를 통해 아세트산, 시트르산, 말산 및 숙신산이 형성됩니다. 고급 알코올은 또한 알라닌으로의 아미노화에 의해 피루브산에서 형성되며, 이는 차례로 상응하는 케토산으로 아미노화됩니다. 알코올 발효 조건에서 케토산은 환원되어 고급 알코올을 형성합니다. 따라서 알코올 발효의 2차 및 부산물을 엄격하게 구분할 수 없습니다.
아세트산 알데히드는 아세트산과 에틸 알코올의 형성과 함께 불균일화를 겪을 수 있습니다(카니자로 반응):
CH 3 COH + CH 3 COH + H 2 O = CH3COOH + CH 3 CH 2 OH.
알데히드 분자 중 하나는 산으로 산화되고 다른 하나는 알코올로 환원됩니다. 알칼리성 환경에서 한 분자
아세트산 알데히드는 아세트 알데히드의 두 번째 분자와 산화 환원 반응을 시작합니다. 이 경우 에틸알코올, 아세트산 및 글리세린이 동시에 형성되며, 이는 다음 총식으로 표현됩니다.
2C 6 Hi 2 0 6 + H 2 0 = 2CH 2 ONSNONCH 2 OH + CH 3 CH 2 OH + CH 3 COOH + 2CO 2.
글리세린은 알코올 발효 과정에서 소량 생성됩니다. 발효 조건이 변경되면 산업적 규모로 생산할 수 있습니다.
글리세린과 아세트알데히드는 알코올 발효의 중간 생성물입니다. 정상적인 발효 과정의 마지막 단계에서 아세트알데히드의 상당 부분이 에탄올로 환원됩니다. 그러나 아세트알데히드가 아황산나트륨과 결합하면 알코올 발효의 방향이 많은 양의 글리세린을 형성하는 방향으로 바뀝니다.
아황산나트륨으로 발효 배지에서 아세트알데히드를 제거하는 방법은 다음과 같습니다.
CH 3 CHO + Na 2 SO 3 + H 2 OW CH 3 CHONaHSO 2 + NaOH.
아황산염과의 결합으로 인해 피루브산의 탈탄산 반응 동안 형성된 아세트산 알데히드는 수소 수용체 역할을 할 수 없습니다. 아세트알데히드의 자리는 포스포디옥시아세톤이 차지하고 환원된 NAD-H 2 로부터 수소를 받아 α-글리세로포스페이트를 형성합니다. 이 반응은 효소 글리세로포스페이트 탈수소효소에 의해 촉매됩니다. 포스파타제의 작용으로 α-글리세로인산은 탈인산화되어 글리세롤로 변합니다. 따라서 Na 2 SO3가 있으면 글리세롤-알데히드 발효가 발생합니다.
C 6 H 12 0 6 = CH3CHO + CH 2 ONSNONCH 2 OH + CO 2.
설탕 아세트알데히드 글리세린
발효 배지에 도입된 아황산나트륨의 양이 증가함에 따라 결합된 알데히드의 양이 상응하게 증가하고 에탄올 및 CO2의 형성이 감소합니다.
산과 아세토인의 형성. 숙신산은 두 분자의 아세트산과 아세트알데히드 한 분자의 탈수소화 및 축합에 의해 형성됩니다(가설 V. 3. Gvaladze 및 Genavois):
2СН 3 С00Н + СН 3 СНО - * С00НСН 2 СН 2 С00Н + СН 3 СН 2 ОН.
알코올 발효 과정에서 숙신산도 글루탐산의 탈아미노화에 의해 형성됩니다. 이 반응에서 수소 수용체는 triosoglycerol al-Dehyde이므로 탈아민 반응은 글리세롤의 동시 축적을 동반합니다.
C 6 Hi 2 0 6 + COOHCH2CH2CHNH2COOH + 2H 2 0 = COHCH 2 CH 2 COOH -b
포도당 글루탐산 숙신산
2CH 2 ONSNONSH 2 OH 3 + NH 3 + CO 2.
글리세린
암모늄은 단백질 합성을 위해 효모에 의해 소비되고 글리세롤과 숙신산은 배지로 방출됩니다.
교육 구연산, Lafon에 따르면, 에서 온다. 아세트알데히드의 9개 분자:
9CH 3 COH + 4H 2 O = (CH 2 COOH) 2 C(OH) COOH + 6CH 3 CH 2 OH.
레몬산
젖산의 형성은 피루브산의 환원으로 설명됩니다.
CH3COOH + H 2 -> CH 3 CH(OH) COOH.
피루브산
그러나 알코올 발효의 중간 생성물인 포스포글리세롤 알데히드의 가수분해 결과로 형성될 가능성이 더 높은 것으로 여겨집니다.
SNOSNONSN 2 OP0 3 H 2 + H 2 0 - * CH 3 CH(OH) COOH + H 3 P0 4.
포스포글리세롤 젖산
알데히드
아세트산과 아세트알데히드의 축합은 아세토인의 형성을 설명합니다.
1) CH3COOH + CH 3 CHO -> - CH3COOCH3 + H 2 O;
디아세틸
2) CH3COCOCH3 + CH3CHO -4 CH3COCONOCH3 + CH3COOH.
디아세틸이 먼저 형성됩니다. 그 다음, 디아세틸의 물에 의한 공액 산화-환원의 불변에 의해 아세트알데히드가 형성되어 아세토인이 형성된다.
아세토인이 환원되면 2,3-부틸렌 글리콜이 형성됩니다.
CH 3 SOSNONSNz + OVER ■ N 2 *± CH 3 SNONSNONSN 3 + OVER.
알코올 발효의 일부 2차 산물의 형성 메커니즘은 아직 완전히 명확하지 않지만 2차 발효 산물의 합성을 위한 주요 출발 물질이 아세트알데히드임에는 의심의 여지가 없습니다.
부산물 중에는 아세트산과 숙신산이 우세하며, 2,3-부틸렌글리콜과 아세트알데히드도 있습니다. 아세토인과 구연산이 아주 소량 존재합니다.
^ 고급 알코올의 형성. 고급 알코올은 알코올 발효의 독특한 부산물입니다. I. Ya-Veselov의 연구에 따르면 발효 중 고급 알코올은 주로 번식기에 발생합니다
누룩. 이 기간 동안 신진 대사의 강도는 아미노기 전환과 탄수화물의 전환 산물에서 케토산 형성과 관련이 있습니다. 재화는 아미노산 사이의 CH(NH) 2 와 CO 라디칼의 교환으로 구성됩니다. 그리고케톤산. 따라서 류신과 피루브산의 알라닌 형성은 다음과 같이 표시됩니다.
(CH 3) 2CHCH 2 CHNH 2 COOH -f CH3COCOOH -> CH 3 CHCH 3 CH 2 COCOOH +
류신 피루빅-포도 이소닐
질산
CH3CHNH2COOH.
이소프로필트르산(유사하게
알코올 발효 방식에서 피루브산) decar-
Boxylated, isovalerian aldehyde로 변환,
이소아밀 알코올로 환원됩니다.
CH 3 CHCH 3 CH 2 고치 -> (CH 3) 2 CHCH 2 CHO - * - CH 3 CHCH 3 CH 2 SON.
이소프로필 포도 이소발레릭 이소아밀
알데히드산알코올
유사한 방식으로, 아밀 알코올은 이소류신으로부터 형성되고, 이소부틸 알코올은 발린으로부터 형성됩니다.
따라서 새로운 아미노산의 합성은 효모 세포에서 탄수화물과 질소 대사 사이의 주요 다리 역할을 하는 피루브산의 참여로 발생합니다.
고급 알코올의 형성은 여러 요인의 영향을 받습니다. 정상적인 발효 온도가 증가하거나 감소함에 따라 고급 알코올의 양은 감소합니다. 발효 배지의 pH를 3에서 5로 적용하면 고급 알코올의 축적이 증가하고 pH가 더 증가하면 감소합니다. 매체의 폭기는 고급 알코올의 합성을 촉진합니다. 폭기된 매체에서 이소부틸 및 이소-아밀 알코올의 함량이 증가합니다. 자당을 함유한 발효 배지에 아미노산을 도입하면 고급 알코올의 축적도 촉진됩니다. 배양 배지에서 퓨젤 오일의 형성은 효모 바이오매스의 축적과 함께 증가합니다. 발효 배지에서 고급 알코올의 함량은 효모의 성장을 억제함으로써 감소될 수 있습니다.
^ 에테르의 형성. 효모 esterases의 참여로 알코올과 산이 관련된 에스테르화 반응이 발생합니다. 일반적으로 에스테르화 반응은 다음과 같이 표시됩니다. RCH 2 OH + RiCOOH -> RCOOCH 2 R! + H 2 0.
따라서 에탄올이 아세트산과 상호 작용하면 에틸 아세테이트(에틸 아세테이트)가 형성됩니다.
C 2 H 5 OH + CH3COOH 5s CH 3 CO 2 C 2 H s + H 2 0.
에스테르의 형성은 이 반응의 성분이 알데히드일 때 더 쉽게 진행됩니다. 알데히드는 쉽게 산화환원 변형을 일으키고 산, 알코올, 에테르를 형성합니다. 이 경우, 알데히드의 모든 변환은 에너지 소비 없이 독립적인 반응으로 수행될 수 있습니다.
따라서 알데가이드로 인해 에테르가 형성될 수 있습니다.
RCHO + HOCRi -> - RCOOCHjRj.
알데히드는 알돌 축합을 겪을 수 있습니다: CH 3 CHO + C "HsCHO = CH 3 CHONCH 2 CHO.
생성된 물질은 알데히드와 히드록실(알코올) 기를 모두 포함합니다.
알코올과 상호 작용할 때 아세트 알데히드는 디에틸 아세탈로 변합니다.
CH 3 CHO + 2C 2 H 5 OH - * CH3CH (OS, H 5) 2 + H 2 0.
설탕의 발효 및 수반되는 모든 과정의 결과로 양조 및 포도주 양조의 맥아즙은 완제품(맥주, 포도주)으로 변합니다. 그 안의 모든 물질은 향과 맛을 결정합니다. 따라서 고급 알코올(프로필, 아밀, 이소아밀, 티로솔, 트립토폴)은 특유의 냄새가 있으며 이미 더 쾌적하고 부드러운 냄새가 나는 에스테르를 제공합니다. 2,3-부틸렌 글리콜. 글리세린은 단맛이 있습니다.
알코올 생산에서 발효 배지는 양조 증류 장치에서 증류하여 알코올을 얻는 성숙한 양조주라고합니다. 발효 중에 형성된 에틸 알코올과 CO2는 세포를 발효 가능한 배지로 외부로 둡니다. 알코올은 발효 가능한 맥아즙에 어떤 비율로든 잘 용해되며 고르게 분포됩니다. 장부! 그것은 먼저 맥아즙에 용해되고 포화되면 기포의 형태로 방출됩니다. 기포 표면에 계면활성 물질(단백질, 펙틴)의 흡착층이 나타납니다. 개별 기포가 서로 붙으면 기포 셀이 생성됩니다. 점차적으로 발효 된 맥아 즙의 표면은 거품으로 덮여 있습니다.
맥주 맥아즙의 발효에는 발아에 의해 전파되는 상부 및 하부 발효 효모가 사용됩니다. 이 효모는 유대류 균류 - 내균류에 속하며 사카로마이세테스 군에 속합니다.
상면 발효 효모를 사용한 맥아즙 발효는 매우 특징적인 외부 특징을 가지고 있습니다. 상단 효모는 발효 중에 위로 올라와 거품의 머리를 형성합니다. 발효가 끝나면 효모의 일부가 통 바닥에 가라앉습니다.
탑 이스트의 발효 능력은 대부분이 라피노스를 전혀 발효시키지 않고 일부 종만 라피노스를 1/3로 발효시킬 수 있다는 점에서 다릅니다.
바닥 효모는 발효 중에 항상 바닥에 가라앉고 발효 맥아즙 표면에 캡을 형성하지 않습니다. 그러나 발효 능력이 최상급 효모와 다릅니다. 라피노스를 완전히 발효시킵니다.
하면 발효 효모는 양조에 가장 널리 사용됩니다. 상면 발효 효모는 주로 어둡거나 특별한 유형의 맥주에 훨씬 덜 자주 사용됩니다.
잉글리시 포터(English porter)와 같은 특별한 유형의 맥주에는 토룰(Torul) 가족(저발효 브리타노이세테스(Brittanoycetes) 속)에 속하는 효모 균류도 사용됩니다. 이 토룰 그룹은 맥주에 특별하고 독특한 풍미를 줍니다. 그들은 발효 초기가 아니라 마지막에 라거 저장고에서 맥주를 숙성시키기 전에 맥주에 도입됩니다.
양조에 사용되는 풀을 먹인 효모는 영양 배지(콜로니)에서의 성장 특성, 세포 크기 및 모양, 당의 발효 정도, 방향족 물질의 형성 등이 다른 인종으로 구분됩니다.
모든 바닥 효모는 포도당, 과당, 자당, 말토스, 라피노스를 발효시키며 이눌린, 유당 및 덱스트린을 발효시키지 않습니다. 상부 효모는 쉽게 포자를 형성하고, 하부 효모는 훨씬 더 어려우며, 맥주 효모의 일부 균주에서는 포자를 얻는 것이 전혀 불가능합니다.
하류 및 상류 효모의 효모 세포는 대부분의 경우 타원형입니다. 세포의 모양은 매우 다양합니다. 셀의 크기도 매우 다양합니다. 따라서 동일한 순수한 효모 배양물에서 더 둥글거나 더 긴 세포를 찾을 수 있습니다.
다양한 효모 배양물의 세포 크기는 세포 직경이 크거나 작을 때 6-8에서 12 마이크론 범위입니다.
양조 효모의 세포에서는 다른 모든 것과 마찬가지로 뚜렷한 막, 다양한 유기물이 포함된 원형질, 크거나 작은 크기의 액포 및 핵이 구별됩니다.
효모 세포의 핵은 일반적으로 특수 염색 기술로 볼 수 있습니다. 그것은 다양한 모양을 가지고 있으며, 싹이 트는 동안 분열하고, 분리된 절반은 신장으로 통과하여 새로운 딸 효모 세포가 형성됩니다. 포자가 형성되는 동안 세포핵은 세포에서 형성되는 포자 수만큼 많은 부분으로 분할됩니다.
세포의 형태학적 구조는 생리학적 상태에 따라 다릅니다. 퇴적물에 있는 굶주린 세포는 과립형 원형질 구조를 가지며 수축된 것처럼 보입니다. 죽은 세포에서 원형질은 막 뒤에 뒤떨어져 있고 세포 자체는 오그라드는 것처럼 보이며 막의 윤곽이 명확하게 불규칙합니다. 이러한 세포는 메틸렌 블루와 같은 염료로 쉽고 강렬하게 염색됩니다. 젊고 새로 싹이 튼 세포는 지방 방울, 볼루틴 및 예비 영양소의 기타 내포물과 같이 빛을 강하게 굴절시키는 더 어두운 내포물이 보이는 일부 장소에서 더 섬세하고 얇은 원형질을 가지고 있습니다. 세포가 노화됨에 따라 더 많은 내포물과 원형질의 더 뚜렷한 입도가 나타나며 액포도 더 눈에 띄고 커집니다.
효모의 생리학적 상태에 따라 세포의 다른 형태학적 구조는 특정 기술로 맥즙 발효의 유리한 조건 또는 불리한 조건을 판단할 수 있는 지표 중 하나입니다.
소련 양조 산업에서 가장 흔한 초원 효모 경주는 776 경주였습니다.
최근에는 RSFSR 발효산업 중앙연구소에서 사육한 11번과 47번 인종도 널리 퍼졌습니다.
바텀 브루어의 효모는 발효 중, 특히 발효가 끝날 때 마치 서로 붙은 것처럼 개별 세포 덩어리로 구성된 플레이크를 형성하는 특성이 있습니다. 이 효모(응집)의 능력은 실제적으로 매우 중요합니다. 맥주 효모의 박편으로 인해 맥주의 빠른 정화가 이루어지며 분리에 의존하지 않고 발효 탱크에서 효모를 수집하고 후속 발효 주기에 재사용할 수 있습니다.
효모의 박편은 효모 껍질의 특성과 세포의 전하에 따라 달라집니다. 수소 이온의 농도가 변하면 쉽게 위반됩니다. 약알칼리성 환경에서는 응집 효모가 분쇄되지만, 특히 산성 환경에서는 효모의 응집이 변화합니다.
가장 큰 응집제 ™ 효모는 pH 4.0-4.4의 배지에서 관찰됩니다. 배지가 산성화되면 효모가 더러워집니다.
발효 온도는 효모 응집의 특성에 큰 영향을 미칩니다. 더 높은 온도에서 응집은 더 낮은 온도에서보다 덜 두드러집니다.
효모의 지방과 밀도도 효모의 침강 속도에 영향을 미치는 것으로 알려져 있습니다. 이와 관련하여 하류 효모는 또한 하류의 가벼운 효모와 유리하게 비교됩니다.
다운스트림 효모의 성장을 위한 최적 온도는 약 25-27°C이고 최소값은 약 2-3°C입니다. 바닥 효모는 낮은 저장 온도를 잘 견딥니다. 이러한 조건에서 장기간 보관하면 발효 에너지에 적응하고 크게 증가합니다.
하면발효 효모는 60~65ºC에서 죽습니다. 이것은 맥주를 저온 살균할 때 중요합니다. 생산 시 최적의 발효 온도는 일반적으로 8~10ºC 사이로 간주되지만 더 높은 온도(10-14 ° C)에서 가장 높은 발효 강도를 얻을 수 있습니다.
많은 물질이 풀뿌리 효모에 대한 강력한 억제 효과가 있습니다. 배지의 에틸알코올 함량이 1.5%를 초과하면 효모의 번식이 느려지고 알코올 함량이 3%를 초과하면 발효능이 현저히 약해집니다. 산은 훨씬 낮은 농도에서 우울하게 작용하며 무기산은 유기산보다 강합니다. 0.5% 농도의 황산은 1-2시간 내에 효모를 죽이며, 유기산 중 1% 농도의 젖산은 효모에 쉽게 견딘다. 아세트산은 효모에 강한 영향을 미칩니다. 0.5% 농도에서는 급격히 억제하고 1% 농도에서는 단시간에 효모를 사멸시킨다.
가장 약한 유기 탄산(H2CO3)도 효모 발달을 억제합니다. 0.2%까지의 배지 농도에서 이산화탄소의 이러한 억제 효과는 주로 효모 싹의 속도를 늦추는 데 영향을 줍니다. 농도가 높을수록 효모의 발효 능력도 억제됩니다. 9-10 atm의 과도한 CO2 압력에서 달성된 1.5%의 이산화탄소 농도는 발효를 중지하지만 효모를 죽이지는 않습니다.
탄산의 억제 효과는 피루브산이 아세트알데히드와 이산화탄소로 분해되는 반응의 억제와 관련이 있습니다(위의 발효 계획 참조).
포르말린과 같은 아세트알데히드는 비교적 낮은 농도에서 효모 세포에 대한 강력한 억제 효과를 갖지만 정상적인 발효 조건에서 발견되는 것보다 높습니다.
퓨젤 오일의 일부인 발효 중 효모에 의해 형성되는 알코올을 포함한 고급 알코올은 효모 세포에 독성이 있습니다. 그러나 발효 맥주에서 발견되는 농도에서는 발효 에너지에 눈에 띄는 영향을 미치지 않습니다. 그러나 발효가 시작되기 전에 배지에 이러한 고급 알코올 농도가 인위적으로 생성되면 효모의 번식 및 발아 과정이 느려지거나 완전히 멈출 수 있습니다.
자외선은 강력하고 효모를 꽤 빨리 죽입니다.
발효 중 맥주의 정화, 발효 정도 및 발효 과정의 속도, 맥주의 향과 맛은 효모의 특성에 따라 달라집니다.
맥주의 향과 맛을 결정하는 능력 측면에서 효모는 화학 분석 방법으로 맥주 구성에서 발생하는 차이를 확립하는 것이 아직 가능하지 않지만 서로 상당히 다릅니다. 이제, 이러한 차이가 화학적으로 결정될 수 없는 효모의 대사 산물에 의존한다는 사실만 알려져 있습니다. 아마도 이러한 산물의 극소량 때문일 것입니다.
맥주 생산을 위해 가장 가치 있는 효모는 맥아즙 추출물을 깊고 빠르게 발효시키고 강한 II 완전한 정화를 제공하며 뚜렷한 향을 형성하고 부드러운 맛맥주.
그러나 효모의 이러한 특성은 어느 한 인종에서 결합되지 않습니다. 따라서 생산에서 그들은 효모의 혼합 인종을 널리 사용하기 시작하거나 다른 인종에서 발효하고 주요 발효 후에 맥주를 혼합합니다. 혼혈의 경우 효모는 동일한 번식률로 선택됩니다. 일부는 발효 능력이 더 뚜렷하고 다른 일부는 향이 더 미묘합니다. 그러나 모든 조건에서 혼합된 효모 균주는 거의 동일한 뚜렷한 응집을 가져야 합니다.
위의 모든 사항은 배양된 효모에 적용됩니다. 그 외에도 생산시 위성 및 해충으로 소위 야생 효모가 있습니다. 그들은 같은 식물 곰팡이 과에 속할 수 있고 사카로마이세테스 그룹과 배양된 맥주 효모에 속할 수 있습니다. 다른 가족의 야생 효모가 종종 생산에서 발견됩니다.
Saccharomycetes에는 효모 Saccharomyces ellі psoideus, Saccharomyces pasteurianus, Saccharomyces apiculatus 등이 포함됩니다. 그들은 발효 에너지가 매우 약하고 1% 이하의 알코올을 형성합니다. 그 중 많은 것들이 이취(곰팡이, 샐러리 냄새)와 맥주 맛의 원인입니다.
생산 해충에는 야생 필름 효모(Mycoderma)도 포함됩니다. 그들은 주로 필름 형태로 맥아 즙과 맥주의 표면에서 발생하고 맥주에 불쾌한 맛과 냄새를주는 물질을 형성하고 알코올은 아세트산과 탄산으로 산화됩니다.
대부분의 야생 효모는 산소를 필요로 하며 10°C 미만의 온도에서는 잘 자라지 않습니다. 따라서 야생 효모의 발생으로부터 생산을 보호하는 예방 조치는 청결 유지, 저온에서 발효, 맥아즙의 접촉면 감소 및 맥주를 공기와 발효시키는 것으로 축소됩니다.

 겨울용 야채 고등어 샐러드 : 길에서, 저녁으로, 점심으로 사무실
겨울용 야채 고등어 샐러드 : 길에서, 저녁으로, 점심으로 사무실 겨울용 야채 통조림 고등어
겨울용 야채 통조림 고등어 팬케이크로 닭고기를 채우는 방법
팬케이크로 닭고기를 채우는 방법 양배추 롤 - 최고의 요리법
양배추 롤 - 최고의 요리법